马上注册,结交更多好友,享用更多功能,让你轻松玩转社区。
您需要 登录 才可以下载或查看,没有账号?注册
×
古神经生物学是通过分析脑内分泌以确定颅内特征和量来研究脑进化的研究。被认为是神经科学的细分,古神经生物学结合了包括古生物学和考古学在内的其他研究领域的技术。它揭示了有关人类进化的具体见解。颅骨的独特之处在于它的生长是响应大脑组织的生长而不是遗传指导而产生的,就像支持运动的骨骼一样。化石的头骨及其内生物可以相互比较,可以与新近死者的头骨和化石进行比较,甚至可以与其他物种的头骨和化石进行比较,从而推断出功能解剖,生理学和系统发育。古神经生物学在很大程度上受到整个神经科学发展的影响。如果没有关于当前功能的大量知识,就不可能推断出古代大脑的功能。[1]
人古生物生物学专门指通过直接检查人类及其近亲人的化石记录(定义为比黑猩猩与人类更近的物种)来研究大脑进化的研究。[2]古神经生物学家分析内分泌物,该内分泌物可复制已印在头骨内表面上的大脑外部形态的细节。[3]

南方古猿
内容
1 历史
1.1 蒂莉·爱丁格
1.2 Holloway与Falk之间的冲突
2 脑内窥镜
3 研究方法
3.1 整体脑容量
3.2 形态分析
3.3 卷积模式与脑组织
3.4 不对称
3.5 脑膜模式
3.6 颅内血管
3.7 相对瓣大小
4 意义
5 局限性
6 感兴趣的研究
6.1 脑的形状,智力和认知能力
6.2 退行性疾病和功能障碍
7 参考
历史

灵长类动物头骨系列
人类对大脑及其功能有着长期的兴趣。关于大脑及其功能的最早记录研究来自于公元前17世纪古埃及人撰写的纸莎草纸文本。该文档详细介绍了48种医疗疾病,并提供了有关如何处理头部伤口的参考。在公元前6世纪晚些时候,古希腊人开始集中精力研究大脑以及视神经与大脑之间的关系。然而,直到人类历史的晚期才开始对大脑进化进行研究。[4]
比较解剖学在19世纪后期开始出现。关于生活的两种主要观点应运而生。理性主义和先验主义。这些构成了这一时期科学家思想的基础。 Georges Cuvier和étienneGeoffroy St. Hilaire是比较解剖学新领域的领导者。 Cuvier相信仅凭经验即可创建功能形态的能力。他强调器官的功能必须与其形式相吻合。相反,Geoffroy则非常重视直觉作为一种理解方法。他的思想基于两个原则:联系原则和计划统一原则。杰弗罗伊是最早在物种间寻求同源性的人之一,尽管他认为这是通用计划的证据,而不是经过修饰的后裔。[4]
19世纪下半叶,在比较解剖学中,受到了查尔斯·达尔文(Charles Darwin)在1859年的《物种起源》中的工作的重大影响。这项工作完全改变了比较解剖学家的观点。在达尔文发布《物种起源》的八年之内,他关于祖先的后裔的观点被广泛接受。这导致了人们试图了解大脑不同部分如何进化的转变。[4]促成古神经生物学的下一个重大创新是显微镜。尽管显微镜是在17世纪发明的,但直到19世纪后期才用于生物学。在显微镜下观察脑细胞的技术花费了很长时间来完善。 1873年,卡米洛·高尔基(Camillo Golgi)手持此工具,开始对大脑进行细胞细化,并采用各种技术来完善轴突显微成像。路德维希·爱丁格(Ludwig Edinger)充分利用了这一点,并提出了一个新的解剖学分支,称为比较神经解剖学。爱丁格认为,脊椎动物是按线性渐进序列进化的。他还认为,大脑的变化是基于一系列的加法和分化,而最高,最复杂的大脑则是最大脑化的大脑。[5] 1885-1935年是比较神经解剖学领域的一个思想爆炸。这个时代的高潮是Arienns,Kappers,Huber和Cosby出版的《神经系统比较解剖学》。这篇论文影响了蒂莉·爱丁格(Tilly Edinger),她后来成为古生物学的创始人。[4]
蒂莉·爱丁格
主条目:蒂莉·爱丁格(Tilly Edinger)
Ottilie“ Tilly” Edinger于1897年出生于德国法兰克福。她的父亲路德维希·爱丁格(Ludwig Edinger)本人是比较神经病学的先驱,为蒂莉(Tilly)的研究领域和整个科学界提供了宝贵的宝贵经验。蒂莉(Tilly)在参加当时唯一的法兰克福女子中学-席勒-舒勒(Schiller-Schule)之前,有很多私人补习老师。蒂莉·爱丁格(Tilly Edinger)继续学习动物学,地质学和古生物学的大学课程。在准备博士学位论文时,爱丁格遇到了中生代海洋爬行动物Nothosaurus的自然脑内铸物。爱丁格的第一篇论文发表于1921年,重点是鼻龙标本的特征。在发表她的著作之前,仅通过对现存鱼类,两栖动物,爬行动物,鸟类和哺乳动物大脑的比较解剖来得出有关脊椎动物大脑进化的推论。蒂莉·爱丁格(Tilly Edinger)在神经病学和古生物学领域的背景为她整合比较解剖学和地层层序铺平了道路,从而将时间概念引入了神经病学并开创了古神经生物学领域。该领域是在1929年出版的《化石大脑》(化石大脑)中正式定义的,该文献汇编了有关该主题的知识,该知识以前散布在各种期刊中,被视为孤立事件。[6]
爱丁格还在德国期间,就开始使用古生物学方法研究现存物种,方法是利用地层学和比较解剖学证据推断海牛的进化性大脑发育。爱丁格在纳粹德国继续她的研究,直到1938年11月9日晚上,成千上万的犹太人被杀或被关押在被称为克里斯塔纳赫特(Kristallnacht)的地方。尽管签证尚未立即提供给移民美国,但在珍视她的工作的朋友和同事的帮助下,爱丁格得以移民到伦敦,在那里她将德语医学文本转移成英语。最终,她的签证配额号码被拨打,她得以移民到美国,在哈佛比较动物学博物馆担任研究员。[6]
她在古神经生物学领域的贡献包括确定内分泌物在多大程度上反映了古代大脑的解剖结构,比较解剖学足以解释大脑的进化,脑内分泌物预测已灭绝生物的生活方式的能力以及大脑的大小是否已增加在地质时期今天仍在探讨的主题。在她的晚年,爱丁格与下一代古生物学家进行了交流,后者确保了她50年职业生涯中的工作持续到未来。她的职业生涯的顶峰成就是1804年至1966年间出版的带注释的古生物学文献参考书目的汇编。书目《古生物学1804-1966》是由同事在1975年因死于爱丁格而过早死于死因的同事完成并出版的。 1967年发生交通事故。[6]
Holloway与Falk之间的冲突
古生物生物学家Ralph L. Holloway和Dean Falk对内生法氏古猿AL 162-28凹陷的解释不同。霍洛威(Holloway)认为,抑郁症是由于在Lambdoid缝合线上打滑而引起的,并且沟纹表明大脑组织正在朝着更人性化的方向运动,而福克(Falk)坚持认为,抑郁症是指月球状沟,这表明猿像沟纹两位科学家之间的争论不仅仅取决于AL 162-28内铸件,还涉及到所有南澳大利亚的化石化石,霍洛韦坚持认为存在人形的沟渠特征,而福尔克则坚持认为这些特征是庞然大物。 Holloway和Falk之间的辩论如此激烈,以至于在1983年至1985年之间,他们发表了四篇关于鉴定Taung内铸物(Australopithecus africanus)的月状沟内侧端的论文,这仅进一步加强了每个科学家各自之间的区分。意见。尽管没有关于这些化石的确切结论,但由于冲突,创造了许多技术或对其进行了严格的分析和完善。这些在内窥分析中的新技术包括使用立体刻绘在不同形状的内窥之间转移苏尔奇,测量来自照片而不是直接来自样本的指标,以及直接从样本和来自照片的测量中混淆测量。[3]
大脑颅腔模型

正常人脑CT扫描
脑内铸是颅骨内部特征的烙印,颅骨捕获了大脑自身在颅骨上施加的压力所产生的细节。内铸可以通过颅骨孔的沉积而自然形成,由于钙的沉积会随着时间的流逝而变得坚硬,也可以人为地通过硅或乳胶制成模具,然后在水浴中放入熟石膏填充模具以平衡力,从而人工形成。保留原始形状。天然内endo非常罕见。大多数研究是人工方法的结果。尽管其名称暗示它是曾经活着的大脑的复制品,但由于被曾经包围并保护大脑组织的软脑膜,蛛网膜和硬脑膜的缓冲作用,内分泌物很少会出现卷积。此外,并非所有内铸物都是从完整的颅骨化石中产生的,随后,根据相似的化石来估算缺失的部分。在某些情况下,来自相同物种的多个化石的碎片被用于构建单个内铸物。[7]
最近,计算机断层扫描在重建内播方面发挥了重要作用。该过程是非侵入性的,其优点是能够在创纪录的时间内分析化石,而几乎没有损坏所审查的化石的风险。 CT成像是通过应用X射线产生断层扫描仪或断层密度图像来实现的,这些图像与MRI扫描过程中产生的图像相似。[8] CT扫描使用约1毫米厚的切片来重建标本的虚拟模型。[9]当化石颅骨被天然内容物占据时,如果不破坏化石的骨骼部分就无法去除,则此方法特别有用。由于颅骨及其内容物的密度不同,因此可以虚拟重建颅内腔及其独特特征。[8]
诸如计算机断层扫描成像或CT扫描之类的射线照相技术,再加上计算机程序设计,早在1906年就已被用于分析脑内投射。[10] 先进的计算机图形技术的最新发展使科学家能够更准确地分析大脑内endo。 华盛顿大学医学院的M. Vannier和G. Conroy开发了一种可对3D表面形貌进行成像和分析的系统。 科学家能够对表面标志进行编码,从而使他们能够分析沟长,皮层不对称性和体积。[11] 美国和欧洲的放射学家,古人类学家,计算机科学家已合作使用虚拟技术研究此类化石。[10]
研究方法

通过MRI观察人脑的矢状面
古神经生物学围绕内分泌的分析。这种分析大部分集中在解释沟渠模式上,这很困难,因为痕迹通常很难辨认,并且没有明确的路标可用作参考点。此外,唯一清晰的参考平面是矢状平面,其特征在于明显的大脑不对称性。由于通常很难从化石细节中获得清晰的数据,因此关于解释的争论很多。经验通常是内播分析中的重要因素。[1]因此,古神经生物学领域的很大一部分来自发展更详细的程序,这些程序增加了解析度和解释的可靠性。
整体脑容量
脑内endo的统计分析可提供有关总体脑容量(“颅内容量”)增加的信息。由于内分泌不是曾经生活过的大脑的精确复制品或精确投射,因此需要计算机算法和CT扫描来计算颅内体积。计算出的颅内体积包括脑膜,脑脊液和颅神经。因此,这些体积最终要大于曾经生活的大脑。[4]此信息对于计算相对大脑大小RBS和脑电商EQ很有用。计算的RBS也必须知道受试者的相应体重。 RBS通过将大脑的重量除以体重来计算。可以根据使用的数据集以几种不同的方式确定EQ。例如,Holloway和Post通过以下公式计算EQ:
在讨论分类学识别,行为复杂性,智力和不同的进化速率的科学文献中,大脑的体积非常重要。在现代人类中,颅骨容量可能相差1000 cc,与行为没有任何关系。这种变化程度几乎等同于从奥古斯丁化石到现代人类的总体积增加,并质疑了依靠颅骨能力作为复杂性的度量是否有效。[12]
许多古生物学家通过浸没法测量颅骨容量,其中将烧杯中的水置换量作为内铸物的体积。认为此方法不够准确的科学家将使用类似的程序,在该程序中,将带有喷嘴的烧杯充满,直到充满为止。然后称量由内浇铸物置换的水,以确定内浇铸物的体积。尽管这两种技术都比以前的方法精确得多,但科学家们对诸如计算机断层扫描之类的更先进的技术将提供更高的体积测量准确性感到乐观。[7]
形态分析
形态分析依赖于内铸表面的弦和圆弧测量。内窥镜的长度,宽度,前reg基底和高度的测量是使用卡尺进行的。[7]额叶,顶叶和枕叶的弦长(沿着中矢面的最宽点处的叶的长度)是使用屈光度计测量的,在该屈光度计中,界标被投影到二维表面上。如果在制作折光仪之前未正确确定内铸件的方向,则测量结果可能会偏斜。经常使用几何形态计量学(叠加在内铸件测量值上的坐标系)来比较不同大小的样本。也可以参考Broca区,以最大长度的25%间隔的内铸件的高度和保险库模块(最大长度,宽度和中间高度的平均值)来进行测量。[13]尽管可以进行其他测量,但研究之间标志性的选择并不总是一致的。[7] [13]
卷积模式与脑组织
卷积是构成大脑褶皱的单个回旋和沟,是内分泌法最难评估的方面。由于覆盖大脑表面的脑膜和脉管系统,大脑表面通常被称为平滑且模糊。如果准确或保留内窥镜,可以观察到潜在的回旋和沟纹模式,但是与这些模式相关的不确定性通常会引起争议。[1]因为健壮的南极奥古斯丁化石显示了这些细节,所以在适当的时候,在内卷积研究中包括了卷积。[7]
不对称
左半球和左半球之间的不对称程度是大多数古生物学家关注的一个问题,因为它可能与标本的惯用性或语言发展有关。由于半球的特殊性而出现不对称,并且以定性和定量方式观察到。半球的不均匀性,被称为花瓣状,其特征在于叶变宽和/或突出超出了对侧叶。例如,惯用右手的人通常比对侧裂片具有更大的左枕叶和右额叶。由于现代人类大脑额叶皮层的交流中心的专长,Petalalias也出现了。枕叶中的花瓣状花序比额叶中的花瓣状花序更易于检测。[7]直立人标本上已记录了某些不对称性,例如180万年前的人redolfensis标本,类似于现代人的不对称性。[4]一些大猩猩显示出很强的花瓣状,但并未像其他人类那样与其他花瓣状结合使用。科学家利用花瓣的存在来显示其复杂性,但它们并不是向人类大脑进化的明确指示。[7]
脑膜模式
尽管脑膜与行为没有联系,但由于物种中脑膜模式的高度保守性,它们仍在古神经生物学领域进行研究,这可能是确定分类和系统发育的一种方法。[7]
颅内血管
由于脑膜血管是大脑最外层的一部分,因此它们经常会在颅腔中留下血管槽,这些血管槽会被内分泌物捕获。颅内血管系统起源于颅骨的孔,并在活体中为颅骨和硬脑膜供应血液。某些化石中的脉管系统保存得很好,因此可以观察到循环系统的末端分支。颅骨脉管系统的分析集中在额叶区域的前脑膜系统,顶颞叶和部分枕前区域的脑膜中部系统以及小脑区域的小脑窝系统。在人的进化过程中,中脑膜系统发生了最大的变化。尽管在上个世纪已经对颅脑血管系统进行了详尽的研究,但由于研究之间结果很少重叠,因此尚未就血管系统分支和模式的识别方案达成共识。因此,颅内血管系统更适合于推断输送到大脑不同部位的血液量。[14]
相对瓣大小
无法通过内铸来确定中央或中央沟的准确位置。仍然可以粗略地了解波瓣的大小。[4]
意义
古神经生物学的研究使研究人员能够检查人类脑病的进化本质。传统上,古神经生物学家一直致力于确定古代大脑的大小以及相关物种中出现的模式。通过找到这些测量值,研究人员已经能够预测物种的平均体重。内播还揭示了古代大脑的特征,包括相对叶大小,血液供应以及对进化物种解剖结构的其他一般见解。[4]
局限性
尽管古生物学在研究脑进化方面很有用,但确实存在对本研究提供的信息的某些限制。化石记录的有限规模和完整性限制了古神经生物学准确记录大脑进化过程的能力。[15]此外,化石的保存对于确保所研究的内铸物的准确性是必要的。[16]风化,侵蚀和整体逐渐变质可能会改变自然恢复的内或由现有化石产生的。[17]大脑的形态也可能难以量化和描述,这使研究的结果更加复杂。[16]此外,古神经生物学对所研究物种的大脑内部的实际解剖结构了解甚少。内分泌的研究仅限于外部解剖。颅内特征之间的关系仍然难以捉摸。比较古神经病学仅揭示了相关物种(例如大猩猩)之间的颅内大小差异。由于大脑的大小和智力之间没有直接的直接关系,因此只能对人类属古代亲属的发育行为做出推断。
古神经生物学的这些局限性目前正在通过开发更先进的工具来完善内分泌研究来解决。
感兴趣的研究
大脑的形状,智力和认知能力
Emiliano Bruner,Manuel Martin-Loechesb,Miguel Burgaletac和Roberto Colomc最近进行的研究研究了矢状中脑形状与心理速度之间的联系。这项研究将人类受试者的认知测试纳入与绝种人类的关系中。他们使用来自102位MRI扫描的年轻成年人的2D图像进行比较。这种相关性很小,表明中矢状脑部几何形状对个体认知表现的影响可以忽略不计,但仍提供了大脑进化特征的有用信息。与顶叶皮层相关的区域似乎参与了大脑几何形状与心理速度之间的关系。[18]
退行性疾病和功能障碍
科学家J. Ghika认为,使用古神经生物学是分析导致帕金森氏病,运动障碍,步态障碍,骨佩吉特病,自主神经失调等疾病的几种神经变性的最佳方法。 Rapoport爵士以前关于智人的阿尔茨海默氏病的一项研究表明,使用达尔文主义的观点更好地了解这种疾病本身及其症状很重要。[19]目的是确定导致局灶性或非对称性脑萎缩的遗传机制,从而导致综合症发作,从而影响步态,手部动作(任何形式的运动),语言,认知,情绪和行为障碍。
另见
Evolution of nervous systems
Paleobiology
Paleoanthropology
Paleontology
参考
Bruner, Emiliano (2003). "Fossil traces of the human thought: paleoneurobiology and the evolution of the genus Homo" (PDF). Journal of Anthropologia Sciences. 81: 29–56.
Bienvenu, Thibaud; Guy, Franck; Coudyzer, Walter; Gillissen, Emmanuel; Roualdes, Georges; Vignaud, Patrick; Brunet, Michel (2011). "Assessing endocranial variations in great apes and humans using 3D data from virtual endocasts". American Journal of Physical Anthropology. 145 (2): 231–236. doi:10.1002/ajpa.21488. PMID 21365614.
Falk, Dean (1987). "Hominid Paleoneurobiology". Annual Review of Anthropology. 16: 13–30. doi:10.1146/annurev.an.16.100187.000305. JSTOR 2155862.
Holloway, Ralph. "Evolution of the Brain in Humans- Paleoneurology" (PDF). Columbia. Retrieved September 27, 2011.
Northcutt, Glen (August 2001). "Changing Views of Brain Evolution". Brain Research Bulletin. 55 (6): 663–674. doi:10.1016/S0361-9230(01)00560-3.
Buchholtz, Emily; Seyfarth, Ernst-August (August 2001). "The Study of "Fossil Brains": Tilly Edinger (1897-1967) and the Beginnings of Paleoneurology" (PDF). BioScience. 51 (8): 674–82. doi:10.1641/0006-3568(2001)051[0674:TSOFBT]2.0.CO;2.
Holloway, Ralph L.; Douglas C. Broadfield; Michael S. Yuan (2004). The Human Fossil Record, Volume Three: Brain Endocasts--The Paleoneurological Evidence. Wiley-Liss. ISBN 978-0-471-41823-8.
Marino, Lori; Uhen, Mark D.; Pyenson, Nicholas D.; Frohlich, Bruno (2003). "Reconstructing Cetacean Brain Evolution Using 计算机断层扫描". The Anatomical Record Part B: The New Anatomist. 272B (1): 107–17. doi:10.1002/ar.b.10018. PMID 12731077.
Poza-Rey, Eva María; Arsuaga, Juan Luis (2009). "Reconstitution 3D par Computerized-tomography (CT) et endocrane virtuel du crane 5 du site de la Sima de Los Huesos (Atapuerca)". L'Anthropologie. 113: 211–21. doi:10.1016/j.anthro.2008.12.004.
Mafart, Bertrand; Gaspard Guipert; Marie-Antoinette de Lumly; Gerard Subsol (17 May 2004). "Three-dimensional computer imaging on hominid fossils: a new step in human evolution studies". Canadian Association of Radiologists Journal. 55 (4): 264–70.
Vannier, M. W.; Conroy, G.C.; Krieg, J.; Falk, D. (1987). "Three-dimensional imaging for primate biology". Proc. Natl. Comput. Graphics Assoc. 3: 156–160.
Holloway, Ralph L (1966). "Cranial Capacity, Neural Reorganization, and Hominid Evolution: A Search for More Suitable Parameters". American Anthropologist. 68 (1): 103–21. doi:10.1525/aa.1966.68.1.02a00090.
Bruner, Emiliano (2004). "Geometric morphometrics and paleoneurology: brain shape evolution in the genus Home". Journal of Human Evolution. 47 (5): 279–303. CiteSeerX 10.1.1.461.2334. doi:10.1016/j.jhevol.2004.03.009. PMID 15530349.
Grimaud-Hervé, Dominique (2004). "Part Five - Endocranial Vasculature". In Holloway, Ralph L.; Broadfield, Douglas C.; Yuan, Michael S. (eds.). The Human Fossil Record, Volume Three: Brain Endocasts--The Paleoneurological Evidence. Wiley-Liss. ISBN 978-0-471-41823-8.
Rogers, Scott W. (2005). "Reconstructing the Behaviors of Extinct Species: An Excursion Into Comparative Paleoneurology". American Journal of Medical Genetics. 134A (4): 349–56. doi:10.1002/ajmg.a.30538. PMID 15759265.
Bruner, Emiliano; Manzi, Giorgio; Arsuaga, Juan Luis (2003). Encephalization and allometric trajectories in the genus Homo: Evidence from the Neandertal and modern lineages (PDF). Proceedings of the National Academy of Sciences of the United States of America. 100. pp. 15335–15340. doi:10.1073/pnas.2536671100. ISBN 978-2-536-67110-7. PMC 307568. PMID 14673084.
Bruner, Emiliano (November 2004). "Geometric morphometrics and paleoneurology: brain shape evolution in the genus Homo" (PDF). Journal of Human Evolution. 47 (5): 279–303. CiteSeerX 10.1.1.461.2334. doi:10.1016/j.jhevol.2004.03.009. PMID 15530349.
Bruner, Emiliano; Manuel Martin-Loeches; Miguel Burgaleta; Roberto Colom (March–April 2011). "Midsagittal brain shape correlation with intelligence and cognitive performance" (PDF). Intelligence. 39 (2–3): 141–147. doi:10.1016/j.intell.2011.02.004.
Rapoport, SI (1988). "Brain evolution and Alzheimer's disease". Review Neurology. Paris. 144 (2): 79–90. PMID 2898165. |

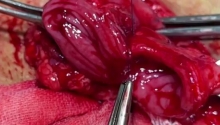
 因缺血而行小肠分段切除及一期吻合术
因缺血而行小肠分段切除及一期吻合术
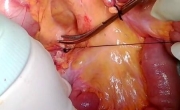
 肌肉活检 _ 如何进行肌肉活检
肌肉活检 _ 如何进行肌肉活检
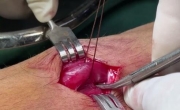
 尿道下裂修复后包皮瘘和龟头隐裂
尿道下裂修复后包皮瘘和龟头隐裂
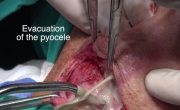 通过外部方法对喉囊肿进行有袋化或去顶
通过外部方法对喉囊肿进行有袋化或去顶

 发表于 2020-2-27 00:01:02
发表于 2020-2-27 00:01:02
 提升卡
提升卡 置顶卡
置顶卡 沉默卡
沉默卡 喧嚣卡
喧嚣卡 变色卡
变色卡 千斤顶
千斤顶 显身卡
显身卡