马上注册,结交更多好友,享用更多功能,让你轻松玩转社区。
您需要 登录 才可以下载或查看,没有账号?注册
×
平衡感是与平衡有关的生理感官之一。 它有助于防止人和动物站立或移动时跌倒。 平衡是许多身体系统共同作用的结果:理想情况下,眼睛(视觉系统),耳朵(前庭系统)和身体在空间中的位置感(本体感觉)是完整的。 前庭系统是三个半规管会聚的内耳区域,与视觉系统协同工作,可在头部移动时使物体保持焦点。 这称为前庭眼反射(VOR)[1]。 平衡系统与视觉和骨骼系统(肌肉和关节及其传感器)配合使用,以保持方向或平衡。 大脑会处理发送到大脑的有关人体相对于周围环境的位置的视觉信号,并将其与前庭和骨骼系统的信息进行比较。

平衡儿童的技能发展

平衡训练
内容
1 前庭系统
2 功能障碍
3 系统概述
3.1 机械
3.2 神经
4 其他动物
5 在植物中
5.1 训练器材
6 参考
前庭系统
主条目:前庭系统

前庭系统图
在前庭系统中,平衡感受力由迷宫中称为内淋巴的液体的液位决定,迷宫中是一组复杂的内耳管。
功能障碍
主条目:平衡失调

该图显示与迷宫性病变引起的旋转诱发的生理性眼球震颤和自发性眼球震颤相关的神经活动。细直箭头-慢速零件的方向;粗直箭头-快速组件的方向;弯曲的箭头-水平半规管中内淋巴的流动方向:AC-前管,PC-后管,HC-水平管。
当平衡感中断时,会引起头晕,迷失方向和恶心。 Ménière病,上管开裂综合征,内耳感染,影响头部的普通感冒或许多其他医疗疾病(包括但不限于眩晕)可能会破坏平衡。快速或长时间加速也可能会暂时干扰它,例如骑旋转木马。打击也会影响平衡接收,尤其是那些在头部侧面或直接在耳朵上的平衡。
大多数宇航员发现,由于他们一直处于失重状态,因此他们在眶上的平衡感受损。这会导致一种晕动病,称为空间适应综合症。
系统概述
 tra.png")
该图线性地(除非另有说明)跟踪所有已知结构的投影,这些投影允许平衡和加速到人脑中的相关端点。
本概述还解释了加速过程,因为其过程相互平衡。
机械
前庭神经支配五个感觉器官。三个半规管(水平SCC,上位SCC,后部SCC)和两个耳石器官(球囊和椭圆囊)。每个半规管(SSC)是一根细管,其厚度在称为骨壶腹e的位置短暂地加倍。在它们的中心底座上都装有一个壶腹式吸盘。吸盘是一种明胶球,与含气小房的立体纤毛相连,受其沐浴的内淋巴的相对运动影响。
由于吸盘是骨迷路的一部分,它会随着实际的头部运动而旋转,并且由于本身没有内淋巴,因此它无法受到刺激,因此无法检测到运动。内淋巴跟随管的旋转,但是,由于惯性,其运动最初滞后于骨迷路的运动。内淋巴的延迟运动会弯曲并激活吸盘。当吸盘弯曲时,连接的立体球与之一起弯曲,激活cr壶腹周围的含气小房中的化学反应,并最终产生由前庭神经携带的动作电位,向身体发出信号,表明其已在空间中移动。
在任何延长的旋转之后,内淋巴会赶上导管,并且吸盘返回其直立位置并复位。但是,当不再继续旋转时,由于惯性,内淋巴会继续弯曲并弯曲,并再次激活吸盘以发出运动变化的信号[2]。
当内淋巴与导管旋转相匹配时,飞行员进行长弯转弯时开始感到直立(不再转弯)。一旦飞行员离开转弯处,便再次刺激了吸盘,从而产生了转弯的感觉,而不是直飞而水平。
HSCC处理绕垂直轴(颈部)的头部旋转,SSCC处理绕横轴的头部移动,PSCC处理绕鼻-尾轴的头部旋转。例如。 HSCC:并排看; SSCC:并肩; PSCC:点头。 SCC发送自适应信号,这不同于两个耳石器官,即球囊和椭圆囊,它们的信号不会随时间而适应。
耳石质膜中刺激纤毛的移位被认为是身体的状态,直到再次刺激纤毛为止。例如。躺着会刺激纤毛,而站起来会刺激纤毛,但是,即使膜恢复了原状,躺着的时间也会保持躺着的信号保持活跃状态。
耳石器质器官具有厚而厚的明胶膜,由于惯性(如内淋巴),其滞后并继续经过覆盖的黄斑,弯曲并激活所包含的纤毛。
椭圆囊对水平面(头到肩)的线性加速度和头部倾斜做出反应,而球囊对垂直面(上下)的线性加速度和头部倾斜做出响应。耳石器器官不动时会更新头部的大脑。运动期间SCC更新。[3] [4] [5] [6]
动纤毛是最长的立体纤毛,位于束中心(每40-70个普通纤毛一个)。如果立体肌向动纤毛运动,去极化会引起更多的神经递质,而与立体肌从动纤毛上倾斜时相比,前庭神经的发射会更多(超极化,神经递质少,发射少)。[7] [8]
神经的
一阶前庭核(VN)项目到IVN,MVN和SVN。
下小脑梗是平衡信息通过的最大中心。这是本体感受和前庭输入之间整合的区域,有助于无意识地保持平衡和姿势。
下橄榄核(也称为橄榄核)通过编码协调时间感官信息来帮助完成复杂的运动任务;这在小脑中被解码并采取了行动。[9]
小脑蚓部有三个主要部分:前庭小脑(由上丘提供的视觉信息和平衡信息进行整合来调节眼部运动),脊髓小脑子(整合视觉,听觉,本体感受和平衡信息以执行身体和肢体运动) 。 (脊髓的)三叉和背柱本体感受输入,中脑,丘脑,网状结构和前庭核(髓质输出)和脑小脑(计划,时间,并在评估了主要来自运动皮层区域的感觉输入后开始运动) ,通过脑桥和小脑的齿状核,输出到丘脑,运动皮层区域和红核)。[10] [11] [12]
绒毛状小叶是一种小脑叶,可通过改变肌肉张力(连续和被动的肌肉收缩)来帮助维持身体平衡。
MVN和IVN位于髓质中,LVN和SVN较小且呈桥状。 SVN,MVN和IVN在姿势纵束(MLF)内上升。 LVN下降至外侧前庭脊髓道内的脊髓,并在骶骨处终止。 MVN也下降到内侧前庭脊髓道内的脊髓,终止于腰椎1。[13] [14]
丘脑网状核将信息分配到其他各种丘脑核,调节信息流。它可以推测性地停止信号,结束不重要信息的传输。丘脑在pons(小脑链接),运动皮层和脑岛之间中继信息。
脑岛也与运动皮层紧密相连。脑岛可能会平衡。
动眼神经复合物是指进入被盖tum(眼动),红核(步态(自然肢体运动)),黑质(奖励)和大脑脚(运动中继)的纤维。卡哈尔核是命名的动眼神经核之一,它们参与眼球运动和反射注视协调。[15] [16]
外展肌只支配眼睛的外侧直肌,使滑车移动眼睛。 Trochlear仅能支配眼睛的上斜肌。滑车和外展肌合拢并放松,以同时将瞳孔指向某个角度并压下眼睛另一侧的球体(例如向下看则将瞳孔向下压并压低(朝向大脑)球体的顶部)。这些肌肉不仅使瞳孔定向,而且经常使它们旋转。 (请参阅视觉系统)
丘脑和上丘通过外侧膝状核连接。上丘(SC)是一种地形图,主要通过视觉输入来实现平衡和快速定向运动。 SC融合了多种感官。[17] [18]

耳朵中流体的流动示意图,继而引起嵌入果冻状吸盘中的含气小房的顶部移位。还显示了负责检测线性加速度或直线运动的椭圆囊和球囊器官。
其他动物
一些动物比人类具有更好的平衡感受力,例如,猫用其内耳和尾巴在细小的栅栏上行走。[19]
在许多海洋动物中,平衡感受器是由完全不同的器官-平衡囊来完成的,平衡囊可以检测微小钙质结石的位置,从而确定哪种方法是“向上”的。
在植物中
主条目:重力
可以说植物表现出一种平衡感受态,即当植物从其正常姿势旋转时,其茎沿向上的方向(远离重力)生长,而其根部向下(沿重力方向)生长,这种现象被称为引力作用,例如杨树茎可以检测到方向和倾斜度。[20]
平衡障碍
自动扶梯损坏的现象
头晕
恩斯特·马赫
意识运动现象
自我运动的幻想
晕车
本体感受
晕船
空间迷失方向
眩晕
训练器材
平衡球
平衡板
平衡自行车
博苏
松弛线
紧绳
单轮车
参考
"Vestibulo-Ocular Reflex - an 概述 | ScienceDirect Topics".
Seeley, R., VanPutte, C., Regan, J., & Russo, A. (2011). Seeley's Anatomy & Physiology (9th ed.). New York, NY: McGraw Hill
Albertine, Kurt. Barron’s Anatomy Flash Cards
"How Does Our Sense of Balance Work?" How Does Our Sense of Balance Work?U.S. National Library of Medicine, 12 Jan. 2012. Web. 28 Mar. 2016.
"半规管s." 半规管s Function, Definition & Anatomy. Healthline Medical Team, 26 Jan. 2015. Web. 28 Mar. 2016.
Tillotson, Joanne. McCann, Stephanie. Kaplan’s Medical Flashcards. Apr. 02. 2013.
Spoor, Fred, and Theodore Garland, Jr. "The Primate 半规管 System and Locomotion." The Primate 半规管 System and Locomotion. 8 May 2007. Web. 28 Mar. 2016.
Sobkowicz, H. M., and S. M. Slapnick. "The 动纤毛 of Auditory 含气小房 and Evidence for Its Morphogenet." Ic Role during the Regeneration of Stereocilia and Cuticular Plates. Sept. 1995. Web. 28 Mar. 2016.
Mathy, Alexandre, and Sara S.N. Ho. "Encoding of Oscillations by Axonal Bursts in 下橄榄核 Neurons." Science Direct. 14 May 2009. Web. 28 Mar. 2016.
Chen, S.H. Annabel, and John E. Desmond. "Cerebrocerebellar Networks during Articulatory Rehearsal and Verbal Working Memory Tasks." Science Direct. 15 Jan. 2005. Web. 28 Mar. 2016.
Barmack, Neil H. "Central Vestibular System: 前庭核 and Posterior 小脑." Science Direct. 15 June 2003. Web. 28 Mar. 2016.
Akiyama, K., and S. Takazawa. "Bilateral Middle Cerebellar Peduncle Infarction Caused by 创伤tic 椎动脉 Dissection." JNeurosci. 01 Mar. 2001. Web. 28 Mar. 2016.
Gdowski, Greg T., and Robert A. McCrea. "Integration of Vestibular and Head Movement Signals in the 前庭核 During Whole-Body Rotation."ARTICLES. 01 July 1999. Web. 28 Mar. 2016.
Roy, Jefferson E., and Kathleen E. Cullen. "Dissociating Self-Generated from Passively Applied Head Motion: Neural Mechanisms in the 前庭核." JNeurosci. 3 Mar. 2004. Web. 28 Mar. 2016.
Takagi, Mineo, and David S. Zee. "Effects of Lesions of the Oculomotor Cerebellar 蚓部 on Eye Movements in Primate: Smooth Pursuit." ARTICLES. 01 Apr. 2000. Web. 28 Mar. 2016.
Klier, Eliana M., and Hongying Wang. "Interstitial Nucleus of Cajal Encodes Three-Dimensional Head Orientations in Fick-Like Coordinates." ARTICLES. 01 Jan. 2007. Web. 28 Mar. 2016.
May, Paul J. "The Mammalian 上丘: Laminar 结构 and Connections." Science Direct. 2006. Web. 28 Mar. 2016.
Corneil, Brian D., and Etienne Olivier. "Neck Muscle Responses to Stimulation of Monkey 上丘. I. Topography and Manipulation of Stimulation Parameters." ARTICLES. 01 Oct. 2002. Web. 28 Mar. 2016.
"Equilibrioception". ScienceDaily. Archived from the original on 18 May 2011. Retrieved 15 January 2011.
Azri, W.; Chambon, C.; Herbette, S. P.; Brunel, N.; Coutand, C.; Leplé, J. C.; Ben Rejeb, I.; Ammar, .; Julien, J. L.; Roeckel-Drevet, P. (2009). "Proteome analysis of apical and basal regions of poplar stems under gravitropic stimulation". Physiologia Plantarum. 136 (2): 193–208. doi:10.1111/j.1399-3054.2009.01230.x. PMID 19453506. |

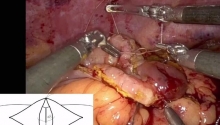
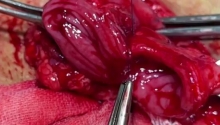
 因缺血而行小肠分段切除及一期吻合术
因缺血而行小肠分段切除及一期吻合术
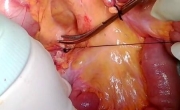
 肌肉活检 _ 如何进行肌肉活检
肌肉活检 _ 如何进行肌肉活检
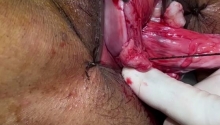
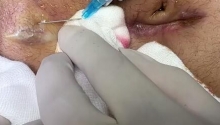
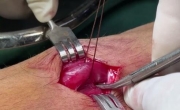
 尿道下裂修复后包皮瘘和龟头隐裂
尿道下裂修复后包皮瘘和龟头隐裂
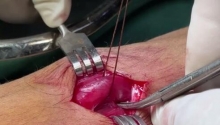
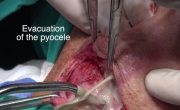 通过外部方法对喉囊肿进行有袋化或去顶
通过外部方法对喉囊肿进行有袋化或去顶

 发表于 2020-1-13 00:00:29
发表于 2020-1-13 00:00:29
 提升卡
提升卡 置顶卡
置顶卡 沉默卡
沉默卡 喧嚣卡
喧嚣卡 变色卡
变色卡 千斤顶
千斤顶 显身卡
显身卡