马上注册,结交更多好友,享用更多功能,让你轻松玩转社区。
您需要 登录 才可以下载或查看,没有账号?注册
×
耳蜗系统是耳蜗降级控制所涉及的听觉系统的组成部分。 它的神经纤维,即少突舌束(OCB),构成前庭尾突神经(第八颅神经,也称为听觉-前庭神经)的一部分,并从脑干的上橄榄复合体(脑桥)伸出到耳蜗。
内容
1 耳蜗系统的解剖
1.1 细胞起源
1.2 脂毛纤维
2 耳蜗系统的生理学
2.1 神经生理学
2.2 电刺激的影响
2.3 MOCS的听觉诱发反应
2.4 MOCS的拟议功能
2.4.1 耳蜗保护装置免受大声声音的侵害
2.4.2 耳蜗功能的开发
2.4.3 噪声中声音的检测和识别
3 参考
耳蜗系统的解剖
细胞源体

哺乳动物的乳突束,分为内侧(红色)和外侧(绿色)系统。两者均包含交叉和未交叉的纤维。主要的纤维由粗线表示。插入物(最左侧)显示了在哺乳动物中观察到的MOCS和LOCS细胞体相对于MSOC和LSOC的位置。
轻齿丛束(OCB)起源于脑干的上橄榄复合体。前庭球囊吻合将传出的轴突带入耳蜗,在那里它们使螺旋器(OC)受神经支配。 OCB包含伸向同侧和对侧耳蜗的纤维,促使最初分为交叉(COCB)和不交叉(UCOCB)系统。[1]然而,最近,OCB的划分基于相对于内侧上橄榄(MSO)的脑干中细胞体的起源部位。腹内侧腹膜腔(MVPO)区,也称为斜方体腹核,位于MSO内侧的神经元弥散区,形成内侧少突舌系统(MOCS)。外侧上橄榄(LSO)是位于MSO外侧的神经元的一个独特核,形成了外侧乳突系统(LOCS)。[2] [3] MOCS神经元是大型的多极细胞,而LOCS的经典定义是由小型球形细胞组成。对于OCB生理学,这种划分被认为更有意义。[4]除了这些经典定义的少突触神经元外,管道追踪方法的进步还有助于揭示第三类绕突触表面的少突触神经元,称为壳神经元。[5]因此,LSO内的LOCS类细胞体被称为内在LOCS神经元,而LSO周围的那些被称为壳或外在LOCS神经元。壳神经元通常很大,在形态上与MOCS神经元非常相似。
疏脂纤维
LOCS(起源于内在神经元和壳神经元)都包含未成纤维状的纤维,这些纤维与投射到内部含气小房的I型螺旋神经节细胞的树突突触。虽然固有的LOCS神经元倾向于较小(直径约为10至15μm),而壳OC神经元则较大(直径约为25μm),但固有的OC神经元具有较大的轴突(与轴突相比,其0.77μm)。壳神经元直径为0.37μm)。相比之下,MOCS包含呈锥形鳞状的神经纤维,可直接支配外部含气小房。[6]尽管LOCS和MOCS都包含交叉的(对侧)和未交叉的(同侧)纤维,但是在大多数哺乳动物物种中,大部分LOCS纤维突出到同侧耳蜗,而大多数MOCS纤维突出到对侧耳蜗[2]。 7] MOCS和LOCS中纤维的比例也因物种而异,但大多数情况下LOCS的纤维数量更多。[8] [9] [10]在人类中,估计有(平均)1,000条LOCS纤维和360条MOCS纤维,[11] [12]然而,个体之间的数量有所不同。 MOCS引起了耳蜗的特定频率的神经支配,因为MOC纤维终止于外蜗中的含气小房,该位置由纤维的特征频率预测到耳蜗中的位置,因此以与原发传入神经元。[6] [13] LOCS的纤维似乎也以异位排列。[14]然而,尚不清楚LOCS纤维的特征频率是否与原发传入神经元的特征频率一致,因为选择性刺激LOCS纤维的尝试在很大程度上没有成功。[15]内源性LOCS衍生的轴突在螺旋器内仅传播约1μm,通常呈螺旋形。他们散发出一簇小的突触弹力,这种弹力的范围很紧凑,通常涉及少于10个IHC。相比之下,壳神经元既在顶端又在基底螺旋,并且可以覆盖螺旋器内的大区域。壳轴突通常覆盖1-2八度音调长度。[16]但是,它们的末端乔木非常稀疏。
耳蜗系统的生理学
神经生理学
耳蜗系统目前所有已知的活性都是通过与钙激活的钾离子通道耦合的烟碱类神经递质受体复合物实现的。这些系统一起对大脑的刺激产生异常的突触反应。耳蜗突触末端包含各种神经递质和神经活性肽。尽管γ-氨基丁酸(GABA)也位于末端,但耳蜗系统采用的主要神经递质是乙酰胆碱(ACh)。乙酰胆碱酯从松果体末端释放会激活一种由烟碱α9[17]和α10亚基组成的进化上古老的胆碱能受体复合物。[18]尽管这些亚基产生了对钙和单价阳离子特别可渗透的配体门离子通道[19],但外部含气小房对ACh活化的细胞反应是超极化的,而不是预期的去极化反应。这是由于相关钾通道的快速激活而引起的。该通道是对罂粟碱敏感的,小电导率的SK2钾通道,它被钙激活,钙很可能通过钙诱导的钙从突触下池中钙储存中的钙释放而释放到细胞质中,作为对烟碱复合体传入钙的响应。[20] ]但是,还不能排除通过烟碱α9α10通道进入的钙也可能直接激活SK通道。因此,在ACh刺激后,从外部含气小房记录的电生理反应显示出较小的内向电流(主要由经由乙酰胆碱受体传入的钙所携带),随后紧随其后的是较大的向外电流(即钾电流),该电流使外毛细胞超极化。
如果在听力发作之前通过手术切除了乳突沟丛,则会损害听觉敏感性。[21]然而,在α9或α10基因的遗传消融后,未观察到这种作用。这可能是由于病变的性质不同所致-手术病变的结果是,所有脂球菌神经支配完全丧失了对含气小房的支配,而遗传操作导致选择性功能丧失-仅针对靶基因。完整的突触末端可释放的任何剩余神经活性物质仍可激活含气小房。确实,通过遗传消融LOCS末端存在的一种神经活性肽后,[22]观察到与手术损伤后相似的结果,表明手术的效果很可能是由于该肽的丢失,而不是突触末端中存在的ACh。
电刺激的影响
在动物中,MOCS的生理学比LOCS的生理学进行了更广泛的研究。这是因为MOCS的髓磷脂纤维更易于电刺激和记录。[15]因此,对LOCS的生理知之甚少。[23]
在体内对动物进行的许多研究都通过放置在神经束上的电极传递的电刺激来刺激了耳蜗束(OCB)。这些研究已经测量了有无OCB刺激的听神经(AN)的输出。 1956年,Galambos通过向第四脑室的地板传递震动刺激(在COCB的讨论下),激活了猫的传出纤维。 Galambos观察到,低强度的点击刺激可以抑制AN的复合动作电位(称为N1电位)。[24]这一基本发现被反复证实(Desmedt和摩纳哥,1961; Fex,1962; Desmedt,1962; Wiederhold,1970)。通过刺激内侧SOC中的MOCS细胞体也观察到了N1的传出抑制,[25]证实N1抑制是MOC(不是LOC)刺激的结果。最近,一些研究人员观察到在刺激中脑中的下丘(IC)期间,对耳蜗神经输出的抑制作用(投射到上橄榄复合体(SOC)上)(Rajan,1990; Mulders and Robertson,2000; Ota)等人,2004; Zhang and Dolan,2006)。 Ota等。 (2004年)还显示,在耳蜗中的N1抑制在与电极在IC中的频率位置相对应的频率处最大,这为传出通路的局部组织提供了进一步的证据。
这些发现导致了目前的认识,即MOC活性会降低OHC的活跃过程,从而导致特定频率的耳蜗增益降低。
MOCS的听觉诱发反应

基本的MOC声反射。听神经对声音做出反应,向耳蜗核发出信号。传入神经纤维从耳蜗核穿过中线到达MOCS的细胞体(位于MSOC附近),MOCS的细胞神经纤维向后伸出耳蜗(红色)。在大多数哺乳动物中,反射的大部分是同侧的(显示为粗线),由交叉的MOCS引起。
脑干中的电刺激可导致(i)刺激整个MOCS;(ii)放电速率(最长400 sec-1)远高于声音所唤起的放电速率(最长60 sec-1),并且( iii)电刺激MOCS纤维以外的神经元。因此,MOCS的电刺激可能无法准确指示其生物学功能,也无法准确显示其作用的自然程度。
MOCS对声音的反应是通过MOC声反射途径(见插图)介导的,该途径先前已使用顺行和逆行标记技术进行了研究(Aschoff等,1988; Robertson和Winter,1988)。内部含气小房的声学刺激向后腹耳蜗核(PVCN)发送神经信号,并且来自PVCN的神经元轴突穿过脑干以支配对侧MOC神经元。在大多数哺乳动物中,MOC神经元主要突出到对侧(形成同侧反射),其余的突出到同侧(形成对侧反射)。
对于纯音,反射的强度最弱,并且随着声音带宽的增加而变强(Berlin等,1993),因此,对于宽带噪声观察到最大的MOCS响应(Guinan等,2003)。研究人员已经测量了用声音刺激MOCS的效果。 Liberman(1989)在猫中发现,对侧声音(导致MOCS刺激)降低了N1电位,这种抑制作用在横切疏脂管束(OCB)后被消除了。在人类中,有关传出动作的最大证据来自声刺激后抑制耳声发射(OAE)。
利用声音刺激激活MOC反射通路,已在豚鼠[13]和猫中[6]记录了单个传出纤维的记录。两项研究均证实,正如Cody和Johnstone(1982)以及Robertson(1984)先前所建议的那样,MOC神经元的频率已急剧调整。他们还表明,MOC神经元的发射率随着声音强度从0 dB到100 dB SPL的增加而增加,并且具有与传入神经元相当的阈值(约15 dB以内)。此外,两项研究均表明,大多数MOC神经元对同侧耳中出现的声音有反应,这与大多数哺乳动物MOC神经元位于对侧时是一致的。[2] [7]尚无人类MOC纤维的记录。因为不可能进行侵入性的体内实验。然而,在其他灵长类动物中,已证明约50-60%的MOC纤维是交叉的(Bodian和Gucer,1980; Thompson和Thompson,1986)。
MOCS的拟议职能
MOCS的假设功能可分为三大类: (i)防止声音过大的耳蜗,(ii)耳蜗功能的发展,以及(iii)检测和辨别噪音中的声音。
耳蜗防止大声响
Cody和Johnstone(1982)以及Rajan和Johnstone(1988a; 1988b)表明,持续不断的声刺激(引起强烈的MOCS反应(Brown等,1998))降低了声创伤的严重性。这种保护作用在存在一种已知能抑制脂疏藻束(OCB)(strychnine)的化学物质的情况下被否定,这暗示了MOCS的作用是保护耳蜗免受大声干扰。 Rajan(1995a)以及Kujawa和Liberman(1997)提供了进一步的证据证明听觉传出具有保护作用。两项研究均表明,如果切断OCB,则由于双耳声音暴露造成的动物听力损失更为严重。 Rajan(1995b)还表明,MOC保护的频率依赖性与耳蜗中MOC纤维的分布大致一致。其他支持MOCS功能的研究表明,MOC刺激可减少与长时间噪声暴露相关的临时阈值偏移(TTS)和永久阈值偏移(PTS)(Handrock and Zeisberg,1982; Rajan,1988b; Reiter and Liberman,1995)。 ,并且MOC反射最强的动物对大声声音的听力损害较小(Maison和Liberman,2000年)。 Kirk和Smith(2003)质疑了MOCS的这种生物学作用,即防止大声声音,他认为实验中使用的声音强度(≥105 dB SPL)在自然界中很少或永远不会发生,因此,如此强度的声音的保护机制不可能发展。 Darrow等人最近质疑了这一主张(MOC介导的耳蜗保护是一种现象)。 (2007年),他认为LOCS具有抗兴奋剂作用,可以间接保护耳蜗免受损害。
耳蜗功能的发展
还存在耳蜗束(OCB)在耳蜗功能发展中的作用的证据。 Liberman(1990)在切断OCB后的6个月内测量了成年猫的单个AN纤维的反应。 Liberman并未发现AN光纤的阈值,调整曲线和I / O功能有任何变化。 Walsh等。 (1998年)进行了类似的实验,但是研究人员切断了新生猫的OCB,并在一年后从AN纤维中记录下来。在没有传入耳蜗的输入的猫中,记录到AN的阈值升高,调谐曲线的锐度降低和SR降低。 Walsh等。 (1998年)提出新生儿去雄激素干扰正常OHC的发育和功能,因此牵涉OCB参与耳蜗活动过程的发展。
检测和辨别噪音中的声音
到目前为止,在静默的实验中(通常在声音衰减的房间或房间中)都观察到了MOC引起的影响。但是,在这种情况下测量耳蜗对声音的响应可能无法揭示MOCS的真正生物学功能,因为进化中的哺乳动物很少处于沉默状态,而MOCS对噪声的响应尤其强(Guinan等,2003)。 Nieder和Nieder(1970a,1970b,1970c)在豚鼠上进行了第一个实验,研究了在有噪声的情况下MOC刺激的影响,他们测量了恒定背景噪声(BGN)中的点击刺激引起的耳蜗输出。在这种情况下,他们发现在MOC刺激期间,单击刺激引起的N1电位增强。使用哺乳动物MOCS的电刺激(Dolan和Nuttall,1988; Winslow和Sachs,1987)和声激活(Kawase等,1993,Kawase和Liberman,1993)都证实了这一发现。温斯洛和萨克斯(Winslow and Sachs,1987)发现刺激OCB:
“ ...使听觉神经纤维能够以较低的信噪比,通过放电速率的变化来发出声调水平的变化信号。”(第2002页)
对这些发现的一种解释是,MOC刺激选择性地降低了听神经对恒定背景噪声的响应,从而使对瞬态声音的响应更大。[15]这样,MOC刺激将减少抑制性和适应性掩蔽的效果,因此,该过程被称为“解掩蔽”或“反掩蔽”(Kawase等,1993; Kawase和Liberman,1993)。 。有人认为抗掩蔽在人类中的发生方式类似(Kawase和Takasaka,1995),并且对选择性聆听有影响,因为MOC激活产生的声音的快速解掩会增加总体信噪比(SNR)。 从而有助于更好地检测目标声音。

来自12位接受前庭神经切除术的受试者的注意过滤器深度,相同的耳朵(三角形)或不同的耳朵(交叉)。显示了组合的平均值(----)和95%的置信区间。在耳蜗束(OCB)病变之后,注意力过滤器深度平均降低了约15%。数据取自Scharf等。 (1997)。[26]
在人类中,以恒定BGN进行的心理物理实验也暗示了寡耳lea束(OCB)与选择性聆听有关。也许与该论文最相关的研究是由沙夫和他的同事们进行的。 1993年,Scharf等人。展示了八名接受单侧前庭神经切除术治疗梅尼埃病的病人的数据,该手术切断了OCB(可能是MOCS和LOCS)。 Scharf等。 (1993年)没有发现受试者手术前后的声调阈值有任何明显的差异。在发现之后不久,Scharf等人。 [26] [27](1994,1997)对总共16例接受了单侧前庭神经切除术的患者进行了一套全面的心理物理实验。[26] [27]他们测量了手术前后在心理听觉任务中的表现,发现在(i)音调检测,(ii)音调强度鉴别,(iii)音调频率鉴别,(iv)响度适应, (v)检出陷波噪声。[26] [27]他们唯一的积极发现是,大多数患者在手术后的耳朵中比在健康的耳朵或手术前的同一只耳朵中发现意外的声音更好。使用截断的探针信号程序获得了该结果,该程序导致患者在每次试验中都期望一定的频率。 12名受试者完成了该实验。[26] [27]他们的程序与Greenberg和Larkin(1968)相似,只是只有50%的试验(不是77%)包含的目标频率与听觉提示频率相同。其他50%的试验包含频率与提示频率不同的探针。另外,仅使用了两个探头频率,一个探头的频率高于目标频率,另一个探头的频率低于目标频率。所有试验均在第一个观察间隔之前包含听觉提示(处于目标频率)。结果用于构建基本的注意力过滤器,该过滤器显示了预期(和提示)目标频率以及两个意外测头频率的检测水平。[26] [27]在两个已发表的报告中(Scharf等,1994,1997),OCB受损的耳朵显示出注意力过滤器,其平均深度比完整的OCB的耳朵低15%-正确。 26] [27]尽管没有办法将这个值经验地转换为dB,但是根据Green和Swets(1966)提出的心理测量函数的粗略估计得出的值为2-3 dB。插图中总结了他们的结果。[26]
Scharf和他的同事认为,在这些病人中切开OCB可以抑制意外频率。 在所有受试者中均未出现这种效果,并且观察到受试者之间的差异很大。 但是,OCB切片后,听觉的其他心理生理特征没有受到影响。 Scharf等。 (26)得出的结论是,OCB介导的耳蜗声音抑制作用是抑制意外声音的原因,因此在正常听力的选择性注意中起着作用。[26] 与Scharf的理论相反,Tan等人。 (2008年)认为OCB在选择性聆听中的作用与暗示或预期音调的增强有关。 这种增强可能是由于MOCS在外部含气小房上的活动导致防遮蔽所致。[28]
尽管Scharf等人(1993年,1994年,1997年)的实验未能在听力的基本心理物理特征(检测意外声音除外)方面没有产生任何明显的差异,但许多其他使用动物和人类的研究都暗示了OCB使用更复杂的刺激来进行噪音监听任务。在恒定的BGN中,已观察到具有完整OCB的恒河猴在元音辨别任务中的表现要优于没有OCB的恒河猴(Dewson,1968)。在猫中,完整的OCB与更好的元音识别(Heinz等,1998),声音定位(May等,2004)和强度区分(May和McQuone,1995)有关。所有这些研究均以恒定BGN进行。在人类中,已经对接受过单侧前庭神经切除术(导致OCB切片)的个体进行了噪声语音识别测量。 Giraud等。 (1997年)观察到在BGN中,健康的耳朵比手术的耳朵在音素识别和语音清晰度方面具有较小的优势。 Scharf等。 (1988)以前研究了听觉注意在语音感知过程中的作用,并建议注意注意力集中在频率区域上有助于语音噪声的辨别。 Zeng等人在2000年报告说,前庭神经切除术并不直接影响纯音阈值或强度歧视,[29]证实了Scharf等人的早期发现。 1994年; 1997. [26] [27]对于噪声听觉任务,他们观察到健康的耳朵与手术的耳朵之间存在许多差异。与May和McQuone(1995)的早期发现一致,在没有输入耳蜗束(OCB)的情况下,观察到噪声中的强度区分稍差。但是,Zeng等人的主要发现与“过冲”效应有关,已发现在手术的耳朵中这种过冲效应显著降低(〜50%)。[29]这种效应最早是由Zwicker(1965)观察到的,其特征是,与在恒定稳态噪声中呈现相比,在噪声开始时呈现的音调检测阈值增加。[30] Zeng等。提出这一发现与MOCS引起的防掩蔽是一致的;就是说,在噪声开始时就没有MOCS引起的抗掩蔽,但是在稳态噪声中却变得活跃了。该理论得到了MOC激活的时间过程的支持; [6] [31]与过冲效应的时间进程相似(Zwicker,1965),[30]以及在感音神经性听力障碍的受试者中,过冲效应受到干扰。损失,MOCS极有可能是无效的(Bacon和Takahashi,1992)。
参考
Rasumssen, G. L. (1960). "Chapter 8: Efferent Fibers of the 耳蜗 Nerve and 耳蜗 Nucleus". In Rasmussen, G. L.; Windle, W. F. (eds.). Neural Mechanisms of the Auditory and Vestibular System. Springfield, IL: Charles C. Thomas. pp. 105–115.
Warr, WB.; Guinan, JJ. (Sep 1979). "Efferent innervation of the 螺旋器: two separate systems". Brain Res. 173 (1): 152–5. doi:10.1016/0006-8993(79)91104-1. PMID 487078.
WARR, W.B.; GUINAN, J.J. Jr.; WHITE, J.S. (1986). Richard A Altschuler; Richard P Bobbin; Douglas W Hoffman (eds.). Organization of the efferent fibers: The lateral and medial Olivocochlea systems. Neurobiology of hearing : the 耳蜗. New York: Raven Press. ISBN 978-0-89004-925-9. OCLC 14243197.
Guinan, JJ.; Warr, WB.; Norris, BE. (Dec 1983). "Differential Olivocochlea projections from lateral versus medial zones of the 上橄榄复合体". J Comp Neurol. 221 (3): 358–70. doi:10.1002/cne.902210310. PMID 6655089.
Vetter, DE.; Mugnaini, E. (Jul 1990). "An evaluation of retrograde tracing 方法 for the identification of chemically distinct 耳蜗 efferent neurons". Arch Ital Biol. 128 (2–4): 331–53. PMID 1702612.
Liberman, MC.; Brown, MC. (1986). "Physiology and anatomy of single Olivocochlea neurons in the cat". Hear Res. 24 (1): 17–36. doi:10.1016/0378-5955(86)90003-1. PMID 3759672.
W. B. Warr (September–October 1980). "Efferent components of the auditory system". The Annals of Otology, Rhinology, and Laryngology. Supplement. 89 (5 Pt 2): 114–120. doi:10.1177/00034894800890S527. PMID 6786165.
Thompson, GC.; Thompson, AM. (Dec 1986). "Olivocochlea neurons in the squirrel monkey brainstem". J Comp Neurol. 254 (2): 246–58. doi:10.1002/cne.902540208. PMID 3540042.
Robertson et al., 1989
Azeredo, WJ.; Kliment, ML.; Morley, BJ.; Relkin, E.; Slepecky, NB.; Sterns, A.; Warr, WB.; Weekly, JM.; Woods, CI. (Aug 1999). "Olivocochlea neurons in the chinchilla: a retrograde fluorescent labelling study". Hear Res. 134 (1–2): 57–70. doi:10.1016/S0378-5955(99)00069-6. PMID 10452376.
Arnesen, AR. (1984). "Fibre population of the vestibulo耳蜗 anastomosis in humans". Acta Otolaryngol. 98 (5–6): 501–18. doi:10.3109/00016488409107591. PMID 6524346.
Arnesen, AR. (1985). "Numerical estimations of 结构s in the 耳蜗 nuclei and 耳蜗 afferents and efferents". Acta Otolaryngol Suppl. 423: 81–4. doi:10.3109/00016488509122916. PMID 3864352.
Robertson, D.; Gummer, M. (1985). "Physiological and morphological characterization of efferent neurones in the guinea pig 耳蜗". Hear Res. 20 (1): 63–77. doi:10.1016/0378-5955(85)90059-0. PMID 2416730.
Robertson, D; Anderson, C.; Cole, K. S. (1987). "Segregation of efferent projections to different turns of the guinea pig 耳蜗". Hearing Research. 25 (1): 69–76. doi:10.1016/0378-5955(87)90080-3. PMID 3804858.
Guinan, John J., Jr (1996). Peter Dallos; Arthur N Popper; Richard R Fay (eds.). The Physiology of Olivocochlea Efferents. The 耳蜗. New York: Springer. pp. 435–502. ISBN 978-0-387-94449-4. OCLC 33243443.
Warr, WB.; Beck, JE.; Neely, ST. (1997). "Efferent innervation of the inner hair cell region: origins and terminations of two lateral Olivocochlea systems". Hear Res. 108 (1): 89–111. doi:10.1016/S0378-5955(97)00044-0. PMID 9213126.
Elgoyhen, AB.; Johnson, DS.; Boulter, J.; Vetter, DE.; Heinemann, S. (Nov 1994). "α 9: an acetylcholine receptor with novel pharmacological properties expressed in rat 耳蜗 含气小房". Cell. 79 (4): 705–15. doi:10.1016/0092-8674(94)90555-X. PMID 7954834.
Elgoyhen, AB.; Vetter, DE.; Katz, E.; Rothlin, CV.; Heinemann, SF.; Boulter, J. (Mar 2001). "α10: a determinant of nicotinic cholinergic receptor function in mammalian vestibular and 耳蜗 mechanosensory 含气小房". Proc Natl Acad Sci U S A. 98 (6): 3501–6. doi:10.1073/pnas.051622798. PMC 30682. PMID 11248107.
Katz, E.; Verbitsky, M.; Rothlin, CV.; Vetter, DE.; Heinemann, SF.; Elgoyhen, AB. (Mar 2000). "High calcium permeability and calcium block of the α9 nicotinic acetylcholine receptor". Hear Res. 141 (1–2): 117–28. doi:10.1016/S0378-5955(99)00214-2. PMID 10713500.
Lioudyno; et al. (2004). "A "Synaptoplasmic Cistern" Mediates Rapid Inhibition of 耳蜗 含气小房". Journal of Neuroscience. 24 (49): 11160–4. doi:10.1523/JNEUROSCI.3674-04.2004. PMID 15590932.
Walsh; et al. (1998). "Long-Term Effects of Sectioning the Olivocochlea Bundle in Neonatal Cats". Journal of Neuroscience. 18 (10): 3859–69. doi:10.1523/JNEUROSCI.18-10-03859.1998. PMID 9570815.
Vetter; et al. (2002). "Urocortin-deficient mice show hearing impairment and increased anxiety-like behavior". Nature Genetics. 31 (4): 363–9. doi:10.1038/ng914. PMID 12091910.
Groff, JA.; Liberman, MC. (Nov 2003). "Modulation of 耳蜗 afferent response by the lateral Olivocochlea system: activation via electrical stimulation of the 下丘" (PDF). J Neurophysiol. 90 (5): 3178–200. doi:10.1152/jn.00537.2003. PMID 14615429.
GALAMBOS, R. (Sep 1956). "Suppression of auditory nerve activity by stimulation of efferent fibers to 耳蜗" (PDF). J Neurophysiol. 19 (5): 424–37. doi:10.1152/jn.1956.19.5.424. PMID 13367873.
Gifford, ML.; Guinan, JJ. (1987). "Effects of electrical stimulation of medial Olivocochlea neurons on ipsilateral and contralateral 耳蜗 responses". Hear Res. 29 (2–3): 179–94. doi:10.1016/0378-5955(87)90166-3. PMID 3624082.
Scharf, B.; Magnan, J.; Chays, A. (Jan 1997). "On the role of the Olivocochlea bundle in hearing: 16 case studies". Hear Res. 103 (1–2): 101–22. doi:10.1016/S0378-5955(96)00168-2. PMID 9007578.
Scharf, B.; Magnan, J.; Collet, L.; Ulmer, E.; Chays, A. (May 1994). "On the role of the Olivocochlea bundle in hearing: a case study". Hear Res. 75 (1–2): 11–26. doi:10.1016/0378-5955(94)90051-5. PMID 8071137.
Tan, MN.; Robertson, D.; Hammond, GR. (Jul 2008). "Separate contributions of enhanced and suppressed sensitivity to the auditory attentional filter". Hearing Research. 241 (1–2): 18–25. doi:10.1016/j.heares.2008.04.003. PMID 18524512.
Zeng, FG.; Martino, KM.; Linthicum, FH.; Soli, SD. (Apr 2000). "Auditory perception in vestibular neurectomy subjects". Hear Res. 142 (1–2): 102–12. doi:10.1016/S0378-5955(00)00011-3. PMID 10748333.
Zwicker, E. (Jul 1965). "TEMPORAL EFFECTS IN SIMULTANEOUS MASKING AND LOUDNESS" (PDF). J Acoust Soc Am. 38: 132–41. doi:10.1121/1.1909588. PMID 14347604.
Backus, BC.; Guinan, JJ. (May 2006). "Time-course of the human medial Olivocochlea reflex". J Acoust Soc Am. 119 (5 Pt 1): 2889–904. doi:10.1121/1.2169918. PMID 16708947. |

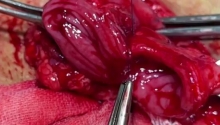
 因缺血而行小肠分段切除及一期吻合术
因缺血而行小肠分段切除及一期吻合术
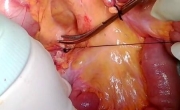
 肌肉活检 _ 如何进行肌肉活检
肌肉活检 _ 如何进行肌肉活检
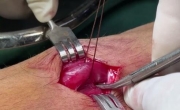
 尿道下裂修复后包皮瘘和龟头隐裂
尿道下裂修复后包皮瘘和龟头隐裂
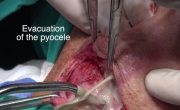 通过外部方法对喉囊肿进行有袋化或去顶
通过外部方法对喉囊肿进行有袋化或去顶

 发表于 2019-12-27 00:00:30
发表于 2019-12-27 00:00:30
 提升卡
提升卡 置顶卡
置顶卡 沉默卡
沉默卡 喧嚣卡
喧嚣卡 变色卡
变色卡 千斤顶
千斤顶 显身卡
显身卡