马上注册,结交更多好友,享用更多功能,让你轻松玩转社区。
您需要 登录 才可以下载或查看,没有账号?注册
×
耳蜗/ kokli /是拉丁语,意为“蜗牛,贝壳或螺丝”,起源于希腊语κοχλαkokhlias。现代定义是内耳的听觉部分,起源于17世纪后期。哺乳动物的耳蜗中存在螺旋器,其中包含含气小房,负责将其从充满液体的周围管道中接收到的振动转化为电脉冲,然后将其发送到大脑以处理声音。[1]据估计,这种螺旋形的耳蜗起源于约1.2亿年前的白垩纪早期。[2]此外,螺旋形耳蜗的听觉神经支配还可以追溯到白垩纪。[3]人类耳蜗的进化是一个重要的科学领域,因为它在化石记录中得到了很好的再现。[4]在上个世纪,许多科学家,例如进化生物学家和古生物学家,竭力开发新的方法和技术,以克服与处理古老而精致的人工制品相关的许多障碍。[2] [5] [6]过去,科学家在完全检查标本而不会造成损坏的能力方面受到限制。[2]在最近的时间里,诸如微CT扫描之类的技术变得可用。[5]这些技术可以在视觉上区分化石动物材料和其他沉积物残骸。[2]通过使用X射线技术,可以确定有关灭绝生物的听觉能力的一些信息,从而洞悉人类祖先及其当代物种。[6]

内容
1 比较解剖
1.1 鳞龙
1.2 恐龙
1.3 哺乳动物
1.4 两栖动物
1.5 鱼
1.6 向现代人类的尼安德特人
2 耳朵
2.1 进化论
2.2 快蛋白的进化
2.3 同质性(趋同进化)
3 参考
比较解剖
尽管鳞翅目(蜥蜴和蛇),弓龙(鸟类和鳄鱼)和哺乳动物的内耳的基本结构相似,并且器官被认为是同源的,但每个组都有独特的听觉器官类型。[2]听力器官在囊状和层状上皮细胞之间的茎状爬行动物的层状导管内产生。在鳞翅类中,听力器官基底乳突通常很小,最多有2000个含气小房,而在始祖龙中,基底乳突可以更长(猫头鹰大于10mm)并且包含更多的含气小房显示了两个典型的极端尺寸,即短和高的含气小房。在哺乳动物中,该结构被称为螺旋器,并显示出含气小房和支持细胞的独特排列方式。科尔蒂(Corti)的所有哺乳动物器官都包含一个由柱状细胞组成的支撑隧道,在其内侧上有内侧含气小房和外侧含气小房。权威的哺乳动物中耳和细长的耳蜗使得对更高频率的灵敏度更高。[2]
鳞甲龙
像在所有的鳞翅类和始祖龙中一样,单小骨(鼻小柱r)的中耳将声音传递到鼻小柱的踏板,从而通过内耳传递压力波。在蛇中,基底乳突大约长1mm,并且仅对低于1 kHz的频率作出反应。相反,蜥蜴倾向于有两个含气小房的区域,一个区域在1 kHz以下响应,另一个区域在1 kHz以上响应。大多数蜥蜴的频率上限约为5–8 kHz。最长的蜥蜴乳头长约2mm,包含2000个含气小房,其传入的神经纤维可以非常尖锐地调谐到频率。[7]
始祖龙
在鸟类和鳄类中,基底乳突的结构相似性背叛了它们的紧密进化关系。基底乳突的最大长度约为10mm,包含多达16500个含气小房。虽然大多数鸟类的听觉上限仅为6 kHz,但the可能听到的声音可达12 kHz,因此接近人类的上限。[8]
哺乳动物
另见:哺乳动物听觉的演变听小骨
产卵的哺乳动物(单足动物(多食的食蚁兽和鸭嘴兽))没有螺旋形的耳蜗,而是一种形状像香蕉的螺旋形,长达约7毫米。像在鳞翅目和始祖龙中一样,它的顶端含有一个lagena(一种前庭感觉上皮)。仅有蜗类哺乳动物(有袋类动物和胎盘类)中的耳蜗真正盘绕了1.5到3.5倍。而在螺旋状单螺旋体中,螺旋器中有许多内部和外部含气小房的行,而在雄性(有袋和胎盘)哺乳动物中,内部毛细胞的行是一排,通常只有三行气小房。[7]
两栖动物
两栖动物具有独特的内耳结构。听力涉及两种感觉乳头,即基底乳头(较高频率)和两栖动物(较低频率)乳头,但尚不确定二者是否与鳞翅目,弓形目和哺乳动物的听觉器官同源,作者不知道它们何时出现。 [9]
鱼
鱼没有专用的听觉上皮,但是使用各种对声音有反应的前庭感觉器官。在大多数硬骨鱼类中,是对声音做出反应的囊状黄斑。在某些鱼类(例如金鱼)中,气囊还具有特殊的骨连接,可增加灵敏度,使听力提高到大约4 kHz。[9]
尼安德特人到现代人类
耳蜗的大小已根据化石记录在整个进化过程中进行了测量。在一项研究中,测量了耳蜗的基础转弯,并假设耳蜗的大小与体重有关。尼安德特人和全新世人类耳蜗基部回旋的大小没有变化,但在早期现代人类和旧石器时代人类中,变大了。此外,在尼安德特人和全新世之间,耳蜗的位置和方向相对于侧渠平面而言是相似的,而早期的现代和旧石器时代的人,其耳蜗的位置要比全新世的人优越。比较中更新世,尼安德特人和全新世的人类时,耳蜗的顶面在人类中的朝向要比后两组低。最后,欧洲中更新世人类的耳蜗比尼安德特人,现代人类和直立人的脸要逊色[10]。人类和猿类是唯一没有高频(> 32 kHz)听力的哺乳动物。[10]人类的耳蜗很长,但是每个频率范围的空间都很大(每倍频程2.5mm),导致频率上限相对降低。[2]人耳蜗绕耳蜗轴(该轴)旋转约2.5圈。[2]像许多哺乳动物和鸟类一样,人类能够感知到仅通过皮皮位移耳膜的听觉信号。[11]
耳朵
由于其在化石记录中的突出地位和保存状态,直到最近,耳朵还被用来确定系统发育史。[4]耳朵本身包含不同的部分,包括外耳,中耳和内耳,所有这些部分都显示出进化变化,而这些变化通常是每种谱系所独有的[14]。正是三叠纪时期鼓膜中耳的独立进化,对陆生脊椎动物各个谱系的听力器官产生了强大的选择压力。[4]
进化论
耳蜗是耳朵的三腔听觉检测部分,由蜗管,鼓阶和前庭阶组成。[9]关于哺乳动物,胎盘和有袋类耳蜗对听觉刺激以及DC静息电位的耳蜗反应相似。[12]这导致人们对这些兽人哺乳动物之间的关系进行了调查,并研究了它们的祖先物种以追踪耳蜗的起源。[2]
有袋和胎盘哺乳动物中都有这种螺旋形的耳蜗,其历史可追溯到大约1.2亿年前。[2]最基本的基底乳突(后来演变为哺乳动物的螺旋器的听觉器官)的发育与脊椎动物的水陆过渡同时发生,大约在3.8亿年前。[7]耳蜗的实际盘绕或螺旋状发生是为了节省头骨内部的空间。[3]耳蜗越长,在相同的听觉范围内,声音频率的潜在分辨率就越高。[3]真正盘绕的哺乳动物耳蜗中最古老的大约长4毫米。[2]
可用于灵长类动物的最早证据显示出短的耳蜗,具有突出的薄片,表明它们具有良好的高频敏感性,而不是低频敏感性。[13]此后,在大约6000万年的时间里,有证据表明灵长类动物发育出更长的耳蜗,而叶片的突出性降低,这意味着它们在低频敏感度方面有所改善,而在高频敏感度方面有所降低。[13]到中新世早期,耳蜗的伸长和片层的退化就完成了。[13]有证据表明,灵长类动物的耳蜗体积与体重之比随时间增加。[3]耳蜗迷宫音量的这些变化会对可听频率的最高和最低产生负面影响,从而导致向下移动。[3]与灵长类动物相比,非灵长类动物的耳蜗迷宫体积总体较小。[3]一些证据还表明,较大的耳蜗迷宫的选择性力可能已在基部灵长类动物结节之后开始。[3]
哺乳动物是许多研究的主题,这不仅是因为要获得有关人类的潜在知识,还因为他们在化石记录中的丰富和丰富的介绍。[14]耳蜗的螺旋形状在哺乳动物的进化途径中演化得比以前认为的要晚,大约在1.2亿年前,在塞里安人分裂成有袋动物和胎盘这两个血统之前。[2]
快蛋白的演变
与耳蜗的进化平行,快蛋白在哺乳动物中显示出更高的进化速率。快蛋白位于哺乳动物耳蜗的含气小房的外部,被认为是运动蛋白。它们存在于包括鱼在内的所有脊椎动物的含气小房中,但最初被认为是膜转运蛋白分子。高浓度的快蛋白仅存在于Therian外含气小房的外侧膜中(单浓度的浓度不确定)。这种高浓度在内部含气小房中找不到,并且在所有非哺乳动物的毛细胞类型中也缺乏。[15]快蛋白在运动性中也有作用,它在陆生脊椎动物的运动功能中起着越来越重要的作用,但是在不同的世系中发展却截然不同。在某些鸟类和哺乳动物中,快蛋白既起转运蛋白的作用,又起着运动的作用,但是向健壮的运动动力的最强进化仅在哺乳动物中进化。据推测,由于螺旋器独特的细胞和骨骼成分,该运动系统在高频下对Therian耳蜗很重要,从而使快蛋白增强整个结构的运动。[2]诸如蝙蝠和齿鲸等现代超声回声定位物种显示出高度进化的快蛋白,并且这些快蛋白随时间显示出相同的序列变化。因此,这些序列显然在不同的时间段内彼此独立地进化。此外,调节外部含气小房的运动反馈的神经递质受体系统(乙酰胆碱)的进化与锡安族人的快蛋白进化相吻合。这表明,在哺乳动物的内耳中,控制系统和运动系统并行发展。[7]
同质性(趋同进化)
陆地脊椎动物在每个主要谱系中独立地进化中耳,因此是平行进化的结果。[7] 因此,一头至尾和哺乳动物的中耳的形态可以解释为趋同进化或同质性。[14] 因此,来自化石的证据证明了从耳朵颌骨脱离的同质性。[14] 此外,很明显,陆地基鼓膜或鼓膜以及诸如喉鼓管之类的连接结构在多种不同的设置下会逐渐演变,而不是一种确定的形态。[16]
参考
Dugdale, DC (2012). "Hearing and the 耳蜗". MedlinePlus.
Manley, GA (August 2012). "Evolutionary Paths to Mammalian 耳蜗e". Journal of the Association for Research Otolaryngology. 13: 733–43. doi:10.1007/s10162-012-0349-9. PMC 3505590.
Luo, Z; Ruf, I; Schultz, JA; Martin, T (July 2010). "Fossil evidence on evolution of inner ear 耳蜗 in Jurassic mammals". Proceedings of the Royal Society B. 282 (1806): 28–34. doi:10.1098/rspb.2010.1148. PMC 2992725.
Clack, JA (November 2002). "Patterns and Processes in the Early Evolution of the Tetrapod Ear". Journal of Neurobiology. 53 (2): 251–64. doi:10.1002/neu.10129.
Ladevèze, S; de Muizon, C; Colbert, M; Smith, T (2010). "3D computational imaging of the petrosal of a new multituberculate mammal from the Late Cretaceous of China and its paleobiologic inferences". Comptes Rendus Palevol. 9 (6): 319–330. doi:10.1016/j.crpv.2010.07.008.
Armstrong, SD; de Bloch, JI; Houde, P; Silcox, MT (2011). "耳蜗 labyrinth volume in euarchontoglirans: implications for the evolution of hearing in primates". The Anatomical Record. 294 (2): 263–266. doi:10.1002/ar.21298.
Manley, GA; Kppl, C (1998). "Phylogenetic development of the 耳蜗 and its innervation". Current Opinion in Neurobiology. 8 (4): 468–474. doi:10.1016/s0959-4388(98)80033-0.
Koeppl, C (2011). "Birds - same thing, but different? Convergent evolution in the avian and mammalian auditory systems provides informative comparative models". Hearing Research. 273 (1): 65–71. doi:10.1016/j.heares.2010.03.095.
Romer, AS; Parsons, TS (1977). "The Vertebrate Body".
Spoor, F; Hublin, J; Braun, M; Zonneveld, F (2002). "The 骨迷路 of Neanderthals". Journal of Human Evolution. 44 (2003): 141–165. doi:10.1016/s0047-2484(02)00166-5.
Drake, R; Vogl, AW; Mitchell, AW (1997). "Gray's anatomy for students". Nature. 390 (6656): 137–142. Bibcode:1997Natur.390..137H. doi:10.1038/36505.
Fernández, C; Schmidt, RS (1963). "The opossum ear and evolution of the coiled 耳蜗". Journal of Comparative Neurology. 121 (1): 151–59. doi:10.1002/cne.901210111.
Coleman, MN; Boyer, DM (2012). "Inner ear evolution in primates through the Cenozoic: implications for the evolution of hearing". The Anatomical Record. 295 (4): 615–631. doi:10.1002/ar.22422.
Luo, ZX (2011). "Developmental patterns in Mesozoic evolution of mammal ears". Annual Review of Ecology, Evolution, and Systematics. 42: 355–380. doi:10.1146/annurev-ecolsys-032511-142302.
Koeppl, C; Forge, A; Manley, GM (2004). "Low density of membrane particles in auditory 含气小房 of lizards and birds suggests an absence of somatic motility". Journal of Comparative Neurology. 479 (1): 149–155. doi:10.1002/cne.20311.
Kitazawa, T; Takechi, M; Hirasawa, T; Adachi, N; Narboux-Name, N; Kume, H; Maeda, K; Hirai, T; Miyagawa-Tomita, S; Kurihara, Y; Hitomi, J; Levi, GL; Kuratani, S; Kurihara, H (2015). "Developmental genetic bases behind the independent origin of the 鼓膜 in mammals and diapsids". Nature Communications. 6 (6853): 6853. Bibcode:2015NatCo...6E6853K. doi:10.1038/ncomms7853. PMC 4423235. PMID 25902370. |

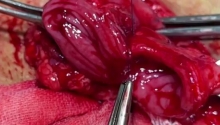
 因缺血而行小肠分段切除及一期吻合术
因缺血而行小肠分段切除及一期吻合术
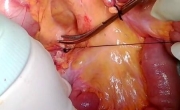
 肌肉活检 _ 如何进行肌肉活检
肌肉活检 _ 如何进行肌肉活检
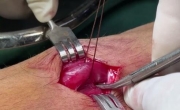
 尿道下裂修复后包皮瘘和龟头隐裂
尿道下裂修复后包皮瘘和龟头隐裂
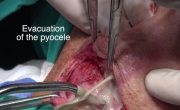 通过外部方法对喉囊肿进行有袋化或去顶
通过外部方法对喉囊肿进行有袋化或去顶

 发表于 2019-12-16 00:00:12
发表于 2019-12-16 00:00:12
 提升卡
提升卡 置顶卡
置顶卡 沉默卡
沉默卡 喧嚣卡
喧嚣卡 变色卡
变色卡 千斤顶
千斤顶 显身卡
显身卡