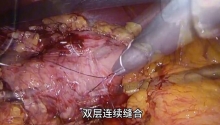
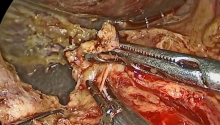
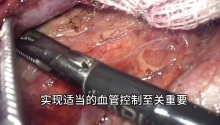
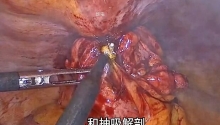
马上注册,结交更多好友,享用更多功能,让你轻松玩转社区。
您需要 登录 才可以下载或查看,没有账号?注册
×
运动控制是具有神经系统的生物体的运动调节。 运动控制包括反射[1]以及定向运动。
为了控制运动,神经系统必须整合多模式感官信息(既来自外部世界也来自本体感受),并发出必要的信号来招募肌肉以实现目标。 该途径跨越了许多学科,包括多感官整合,信号处理,协调,生物力学和认知[2] [3]。 成功的运动控制对于与世界互动以实现目标以及调节平衡与稳定至关重要。
内容
1 感觉运动反馈
1.1 对刺激的反应
1.2 闭环控制
1.3 开环控制
2 协调
2.1 反射
2.2 协同作用
2.3 运动程序
2.4 冗余
3 运动控制中的感知
3.1 基于模型的控制策略
3.1.1 推理和间接感知
3.1.2 正向模型
3.1.3 逆模型
3.2 基于信息的控制
3.2.1 直接感知
3.2.2 行为动力学
4 生理基础
4.1 运动单元
4.2 机理与结构
4.2.1 低阈值和高阈值
4.2.2 顺序
5 参考
感觉运动反馈
对刺激的反应
觉察到感觉刺激并使用该信息影响动作的过程分阶段进行。简单任务的反应时间可以用来揭示有关这些阶段的信息。反应时间是指从出现刺激到反应结束之间的时间段。运动时间是完成运动所需的时间。最初的反应时间实验是由弗朗西斯·唐德斯(Franciscus Donders)进行的,他们利用选择任务的响应时间差异来确定处理刺激并选择正确响应所需的时间。[4]尽管这种方法最终有缺陷,但它产生了这样的想法,即反应时间由刺激识别,随后的反应选择组成,最终最终完成正确的运动。进一步的研究提供了这些阶段确实存在的证据,但是任何反应时间的响应选择期都会随着可用选择数量的增加而增加,这种关系被称为希克定律。[5]
闭环控制
人体运动的闭环系统的经典定义来自Jack A. Adams(1971):[6]。通过错误检测机制,使用反馈将所需输出的参考与实际输出进行比较,从而对错误进行校正。在日常活动中执行的大多数运动都是通过访问感官信息并使用它来更准确地继续运动的连续过程形成的。这种类型的运动控制称为反馈控制,因为它依靠感觉反馈来控制运动。反馈控制是运动控制的一种形式,它依赖于有关性能的感觉信息以及来自执行运动的环境的特定感觉输入。在处理过程中,这种感觉输入并不一定会引起对动作的自觉意识。闭环控制[7]是一种基于反馈的运动控制机制,其中对环境的任何行为都会产生某种变化,这种变化会通过反馈影响未来的表现。闭环运动控制最适合于连续控制的动作,但是对于弹道动作而言,动作速度不够快。弹道动作是指即使没有必要再继续进行下去而无需考虑的动作。由于反馈控制依赖于感官信息,因此其速度与感官处理一样慢。这些运动受速度/精度折衷的影响,因为使用感官处理来控制运动,所以运动进行得越快,其准确性就越差。
开环控制
Jack.A.的经典定义。 Adams是:[8]“开环系统没有反馈或错误调节机制。系统的输入事件会产生影响,系统会对输入进行转换,系统会产生输出……固定时间的交通信号灯会在负载沉重时阻塞交通,并在交通拥堵时阻碍交通很轻。该系统没有补偿能力。”
但是,某些运动发生得太快而无法整合感官信息,因此必须依靠前馈控制。开环控制是运动控制的前馈形式,用于控制快速弹道运动,这种运动在处理任何感官信息之前就结束了。为了最好地研究这种控制方式,大多数研究都集中在脱除咖啡因的研究上,通常涉及猫或猴子的感觉神经与脊髓断开。那些从手臂上丢失了所有感官信息的猴子在从脱除果蝇过程中恢复后恢复了正常行为。大部分技能都可以重新学习,但是精细的运动控制变得非常困难。[9]已经表明,开环控制可以适应不同的疾病状况,因此可以通过改变控制系统的成本功能来用于提取不同运动障碍的特征。[10]
协调
运动控制的核心问题是协调电机系统的各个组件以协调一致地产生运动。电机系统非常复杂,由许多不同组织级别的许多相互影响的部分组成
周围神经元接收来自中枢神经系统的输入并支配肌肉。继而,肌肉产生可驱动关节的力。对于电机系统来说,如何使各个部件协同工作是一个具有挑战性的问题,如何解决此问题是运动控制研究的活跃领域。
反射
在某些情况下,运动组件的协调是硬连线的,由固定的神经肌肉通路(称为反射)组成。反射通常以自动和固定的运动响应为特征,它们发生的时间比依赖于感知过程的反应要快得多。[11]反射在稳定电机系统,为小的扰动提供几乎立即的补偿并保持固定的执行模式方面起着基本作用。一些反射环仅通过脊髓布线,而没有从大脑接收输入,因此不需要注意或有意识的控制。其他人涉及较低的大脑区域,可能会受到事先的指示或意图的影响,但它们仍独立于感知过程和在线控制。
最简单的反射是单突触反射或短环反射,例如单突触拉伸反应。在该示例中,当传入神经元由于肌肉的拉伸而变形时,它们被肌肉纺锤体激活。这些传入神经元在脊髓中直接突触调节相同肌肉收缩的α运动神经元。[12]因此,肌肉的任何拉伸都会自动发出该肌肉反射性收缩的信号,而无需任何中央控制。顾名思义,它的含义是,单突触反射依赖于传入感觉神经元和传出运动神经元之间的单个突触连接。通常,单突触反射的动作是固定的,不能被意图或指令控制或影响。但是,有一些证据表明,这些反射的增益或大小可以根据上下文和经验进行调整。[13]
多突触反射或长环反射是反射弧,其在脊髓中涉及多个单突触连接。这些循环可能还包括大脑的皮质区域,因此由于行程时间较长,因此比其单突触对应区域慢。然而,由多突触反射环控制的动作仍比需要感知处理的动作要快。[14]虽然短环反射的动作是固定的,但多突触反射通常可以通过指导或先前的经验来调节。[15]长环反射的常见示例是婴儿中观察到的不对称的强直性颈部反射。
协同作用
运动协同作用是多元素系统的神经组织,其(1)组织一组元素变量之间的任务共享; (2)确保基本变量之间的协变量,以稳定性能变量。[16] [17]协同作用的组件无需物理连接,而是通过它们对有关正在执行的特定运动任务的感知信息的响应进行连接。协同作用是学习的,而不是像反射那样被硬连线,而是以任务相关的方式组织的;协同作用是针对特定动作而构建的,而对于组件本身通常并不确定。尼古拉·伯恩斯坦(Nikolai Bernstein)在专业铁匠的锤击动作中展示了协同作用。控制锤子运动的手臂肌肉以信息的方式链接在一起,从而使一只肌肉中的错误和可变性通过另一只肌肉的动作自动得到补偿。这些补偿性动作具有反射性,因为它们的发生速度快于感觉处理所允许的速度,但是它们仅出现在专家级演奏中,而没有出现在新手中。对于铁匠,所讨论的协同作用是专门为锤击动作而组织的,不是手臂肌肉的通用组织。协同作用除了依赖于任务外,还具有两个定义特征。共享和灵活性/稳定性。[18]
“共享”要求特定运动任务的执行取决于构成协同作用的所有组件的组合动作。通常,涉及的组件比特定任务所需的组件要严格得多(请参见下面的“冗余”),但是该运动任务的控制仍然分布在所有组件中。一个简单的演示来自两个手指的力量生产任务,在该任务中,要求参与者通过用两个不同的手指按下两个力板上来产生固定量的力量。[19]在此任务中,参与者通过组合独立手指的贡献来产生特定的力输出。尽管任何一个手指产生的力都可以变化,但这种变化会受到另一根手指的作用的限制,从而始终产生所需的力。
协变量还为运动任务提供“灵活性和稳定性”。再次考虑力的产生,如果一个手指没有产生足够的力,则可以用另一根手指来弥补。[19]运动协同作用的组成部分有望改变其作用,以补偿可能影响运动任务结果的其他组成部分的错误和可变性。这提供了灵活性,因为它允许针对特定任务的多种电动机解决方案,并且通过防止单个电动机组件中的错误影响任务本身来提供电动机稳定性。
协同作用简化了运动控制的计算难度。协调人体中众多自由度是一个具有挑战性的问题,这既是由于运动系统的巨大复杂性,也是由于这种组织可能发生的不同水平(神经,肌肉,运动,空间等)。由于协同作用的各个组件在功能上耦合用于特定任务,因此可以通过使用单个神经信号激活相关协同作用来完成运动任务的执行。[20]由于组织由于组件的系统协变而自动出现,因此消除了独立控制所有相关组件的需要。类似于反射是物理连接的,因此不需要中枢神经系统控制单个组件,由于动作是功能连接的,因此可以通过协同作用以最小的执行控制来执行动作。除了运动协同作用外,最近还引入了感觉协同作用这一术语。[21]据信感觉协同作用在整合环境输入的混合物以向CNS提供低维信息从而指导运动协同作用的募集方面起着重要作用。
运动程序
协同作用表示协调是从运动成分的外围相互作用中获得的,而运动程序是特定的,预先构造的运动激活模式,由中央控制器(在生物体的情况下为大脑)生成并执行。[14]它们代表了自上而下的运动协调方法,而不是协同作用提供的自下而上的方法。运动程序以开环方式执行,尽管感觉信息最有可能用于感测生物体的当前状态并确定适当的目标。但是,一旦执行了程序,就无法通过其他感官信息在线对其进行更改。
运动程序的存在的证据来自对快速运动执行的研究以及一旦开始运动就改变这些运动相关的困难。例如,被要求快速摆臂的人在开始运动后被提供“停止”信号时,很难停止该运动。[22]即使在初始“ GO”信号之后但在实际开始运动之前出现停止信号,这种反转困难仍然存在。这项研究表明,一旦开始选择和执行电机程序,它必须运行到完成才能采取其他措施。即使完全阻止了由特定电机程序执行的运动,也可以发现这种效果。尝试执行特定动作(例如用手臂推动)但在不知不觉中已阻止其身体动作的人,在实际发生任何动作之前,他们会显示出相同的肌肉激活模式(包括稳定和支持性激活)动作),就像允许他们完成其预期的动作一样。[23]
尽管有关运动程序的证据似乎很有说服力,但对该理论有一些重要的批评。首先是存储问题。如果一个生物体能够产生的每个运动都需要自己的运动程序,那么对于那个生物体来说,似乎有必要拥有无限数量的此类程序库,并且将这些变量保存在何处尚不清楚。除了这种设施需要大量的存储空间外,还没有发现大脑中的运动程序存储区域。第二个问题与运动的新颖性有关。如果任何特定的运动需要特定的运动程序,则不清楚如何产生一种新颖的运动。充其量,一个人必须在成功执行任何新动作之前先进行练习,而在最坏的情况下,由于没有针对新动作的运动程序,因此将无法进行新动作。这些困难导致了运动程序概念的细微差别,即广义运动程序。[14]通用运动程序是针对特定动作类而不是特定运动的程序。该程序通过环境和生物体的当前状态进行参数设置。
冗余
协调运动系统的一个重要问题是运动自由度的冗余问题。如“协同作用”部分中所述,许多动作和动作可以多种方式执行,因为控制这些动作的功能协同作用可以在不改变动作结果的情况下发生变化。这是可能的,因为与动作的物理约束相比,动作的生成所涉及的运动组件更多。例如,人的手臂有七个关节,这些关节决定了手在世界上的位置。但是,仅需要三个空间尺寸即可指定手可以放置的任何位置。运动学上的自由度过大意味着存在与手的任何特定位置相对应的多个手臂配置。
关于运动冗余性研究的一些最早且最具影响力的工作来自俄罗斯生理学家尼古拉·伯恩斯坦(Nikolai Bernstein)。伯恩斯坦(Bernstein)的研究主要关注了解如何为熟练的动作开发协调。他观察到,电动机系统的冗余性使得有可能以多种不同的方式执行动作和运动,同时获得相同的结果。[20]这种等效的动作意味着在期望的动作与执行这些动作所需的运动系统的协调之间不存在一一对应的关系。任何所需的运动或动作都不具有使之成为可能的神经元,肌肉和运动学的特殊协调性。该电动机等效问题被称为自由度问题,因为它是电动机系统中具有可用冗余冗余度的产品。
运动控制感知
与感知信息的处理如何影响动作和动作的控制相关但又不同的是,对世界结构动作的感知如何的问题。感知在运动控制中非常重要,因为它包含有关对象,环境和身体的相关信息,这些信息用于组织和执行动作和动作。感知以及随后的信息如何用于组织电机系统是当前和正在进行的研究领域。
基于模型的控制策略
大多数基于模型的运动控制策略都依赖于感知信息,但假定该信息并不总是有用的,垂直的或恒定的。眨眼会中断光学信息,环境中的物体会阻碍运动,扭曲会改变物体形状的外观。基于模型和重新引入的控制策略是指那些依赖于环境的准确内部模型的策略,这些模型是由感知信息和先验知识的组合构成的,即使在没有感知信息的情况下,也可以作为计划和执行动作的主要信息。 [24]
推理与间接感知
感知系统的许多模型都采用间接感知,或者认为被感知的世界与实际环境不同的概念。环境信息在被感知之前必须经历几个阶段,并且这些阶段之间的过渡会带来歧义。实际被感知的是根据先前的经验,大脑对环境中正在发生的事情的最佳猜测。支持这种想法的是埃姆斯房间的错觉,其中扭曲的房间使观看者看到的物体在房间内移动时会随着大小的增长或缩小而不断变化。房间本身被视为正方形,或至少由直角组成,因为感知器遇到的所有先前房间都具有这些属性。这种歧义的另一个例子来自特定神经能量学说。该学说提出的发现是,对于不同类型的感觉输入,存在截然不同的神经类型,并且这些神经无论刺激方法如何均以特征性方式做出反应。也就是说,红色导致视神经以特定的模式发射,这种模式由大脑在经历红色时处理。但是,如果以相同的模式对同一条神经进行电刺激,则当不存在相应的刺激时,大脑可能会感知为红色。
正向模型
正向模型是运动控制的预测性内部模型,该模型采用可用的感知信息并结合特定的运动程序,并尝试预测计划的运动运动的结果。前向模型通过确定电机组件的力,速度和位置如何影响环境和个人的变化来构造动作。有人提出,当个体与其环境互动时,正向模型有助于神经控制肢体僵硬。人们认为前向模型使用运动程序作为输入来预测动作的结果。当前向模型的预测与运动的实际结果不匹配时,会生成错误信号,从而提示更新现有模型并提供学习机制。这些模型解释了为什么无法为自己挠痒痒。当感觉无法预测时,感觉会有些发痒。但是,前向模型可以预测您运动的结果,这意味着运动是可以预测的,因此不会发痒。[25]
前向模型的证据来自运动适应性研究。当人的目标指向的到达动作受到力场的干扰时,它们会逐渐但稳定地调整手臂的动作,使他们再次达到目标。但是,它们这样做的方式保留了一些高级运动特性。钟形速度曲线,手的直线平移以及平稳连续的运动。[26]尽管这些运动特征需要惊人的不同的手臂动力学(即扭矩和力),但仍可以恢复。这种恢复提供的证据表明,促使运动的是特定的运动计划,并且该人正在使用正向模型来预测手臂动力学如何改变手臂的运动以实现特定的任务级别特征。预期手臂运动与观察到手臂运动之间的差异会产生一个误差信号,该误差信号将用作学习的基础。前向模型的其他证据来自于实验,实验要求受试者在不可见的运动之后确定效应器的位置[27]。
逆模型
逆模型可预测运动组件的必要运动,以实现所需的感知结果。他们还可以获取运动的结果,并尝试确定导致该状态的运动命令的顺序。这些类型的模型对于开环控制特别有用,并且允许特定类型的运动,例如在头部移动时固定在固定的对象上。作为正向模型的补充,反向模型试图估计如何实现特定的感知结果,以生成适当的运动计划。由于反向模型和正向模型是如此紧密地联系在一起,因此内部模型的研究通常被用作两种模型类型在作用中的证据。
因此,运动适应性研究也为逆模型提供了依据。运动运动似乎遵循预定义的“计划”,该计划保留了运动的某些不变特征。在上述的到达任务中,钟形速度分布和平滑,笔直的轨迹的持续存在为此类计划的存在提供了证据。[26]通过逆模型估计实现这些期望的任务级结果的运动。因此,适应过程是通过以下过程进行的:使用逆模型估计必要的运动,使用前向模型模拟那些运动计划的结果,观察所需结果与实际结果之间的差异以及为将来的尝试更新模型。
基于信息的控制
基于模型的控制的替代方法是基于信息的控制。信息控制策略根据有关环境的感知信息来组织动作和动作,而不是基于认知模型或对世界的再现。电机系统的动作是通过有关环境的信息和有关代理的当前状态的信息来组织的。[28]基于信息的控制策略通常将环境和有机体视为一个单独的系统,而行动是该系统相互作用的自然结果。基于信息的控制策略的核心假设是,对环境的感知包含丰富的信息,并且对于采取行动的目的而言,这是真实的。这与基于模型的控制策略对间接感知的假设背道而驰。
直接感知
认知意义上的直接感知与中殿或直接现实主义的哲学概念相关,因为它基于以下假设:作者感知的是世界上实际存在的东西。詹姆斯·吉布森(James J. Gibson)被誉为将直接感知重塑为生态感知。[29]虽然间接感知的问题表明由于感官信息的含糊不清,所以作者无法获得有关环境中物体的物理信息,但直接感知的支持者(如Gibson)建议,编码在感官信号中的相关信息不是物体的物理特性,而是环境提供的行动机会。这些能力可以直接理解而没有歧义,因此排除了对内部模型或世界重塑的需求。支付仅作为代理与其环境之间相互作用的副产品而存在,因此感知是一种“生态”努力,取决于整个代理/环境系统,而不是孤立的代理。
由于能力是行动的可能性,因此感知与行动和动作的产生直接相关。感知的作用是提供指定应如何组织和控制动作的信息,[30]并且“调整”电动机系统以特定方式响应特定类型的信息。通过这种关系,环境信息决定了电机系统的控制和动作的执行。例如,门口“ affords”通过,但墙壁没有。从环境接收到的视觉信息,以及感知到的关于自己身体的信息,指定了一个人如何通过门口。这些信息共同决定了门口的通行能力,而不是墙壁。另外,朝向并经过门口的动作会产生更多的信息,这反过来又指明了进一步的动作。直接感知的结论是,动作和感知是至关重要的联系,没有一个就无法完全理解。
行为动力学
行为控制理论建立在直接感知行为动力学的假设之上,该行为控制理论将感知有机体视为以功能性方式对信息变量做出反应的动态系统。[28]在对行为的这种理解下,行动是有机体和与环境有关的可用信息之间相互作用的自然结果,这些信息在与身体相关的变量中指定。行为动力学的许多研究都集中在运动上,其中视觉指定的信息(例如光流,接触时间,光膨胀等)用于确定如何导航环境[31] [32]相互作用人与环境之间的力也会影响行为动力学,如肢体僵硬的神经控制所见。
生理基础
运动单元
日常任务,例如步行去洗手间,与您的一个朋友聊天或吃晚饭,都需要多条肌肉来支配身体的各个部位,以正确地移动身体以完成特定任务。由数十,数百甚至数千个运动神经分支组成的运动单元与肌肉相连。在作者的身体中,股直肌含有大约一百万条肌肉纤维,由大约1000条运动神经控制。在一个运动单元(可以分为I型(慢肌)或II型纤维(快肌))内,肌肉纤维的组成类型是一致的(均质)。但是,在一块肌肉中,是两种运动单位(异质)的几种不同组合。[33]
肌肉纤维主要分为三种类型:I型,IIa型和IIb型。如上所述,I型肌纤维被称为慢肌氧化,IIa是快肌氧化,IIb是快肌糖酵解。这三种不同类型的纤维专门具有独特的功能。 I型纤维被描述为高耐久性,但力/功率/速度产量低,IIb型纤维被描述为低耐久性,但力/功率/速度产量高,而IIa型纤维则介于两者之间。

神经系统组织
运动单位是捆绑在一起的多条肌纤维。当一个人想要移动自己的身体来完成某项任务时,大脑会发送一个脉冲信号,该信号通过脊髓到达特定的运动单元。在从大脑接收到信号后,运动单元收缩组内的肌肉纤维以产生运动。动力单元没有部分发火,这意味着一旦检测到信号,该单元内的所有肌肉纤维就会收缩。但是,强度不同。由于每个运动单元一旦受到刺激就会收缩100%的纤维,因此产生不同力或速度的运动单元的类型非常重要。
低阈值和高阈值
对于低强度任务,使用具有较少肌纤维的较小运动单元。这些较小的运动单元称为低阈值运动单元。它们由I型纤维组成,收缩速度要慢得多,因此为日常基本运动(例如在键盘上打字)提供的作用力较小。对于更复杂的任务,使用包含II型肌纤维的运动单元。这些快速抽搐运动单元被称为高阈值运动单元。低阈值运动单元(慢速抽搐运动单元)和高阈值运动单元(快速抽搐运动单元)之间的主要区别在于,与低阈值运动单元相比,高阈值运动单元控制更多的肌纤维并包含更大的肌纤维。另一方面,慢肌纤维(I型)和快肌纤维(II型)的主要区别在于尺寸偏差的理论相同。[34]
招募顺序
在举起重物(例如用哑铃锻炼)的过程中,不仅要抓起低阈值的运动单元,还要招募高阈值的运动单元,以补偿除了握住叉子外还需要的力,其中产生的能量低阈值运动单元足以完成这项工作。进行工作时,身体首先会募集慢速抽搐的运动单元,然后随着完成动作所需的力的增加,会募集更多的更快速抽搐的运动单元。因此,当身体必须携带一个非常重的物体时,它将招募所有可用的运动单位来收缩已使用的特定肌肉。这种现象由Elwood Henneman描述,被称为Henneman的尺寸原理。
纤维与神经
纤维的类型(I型与II型)由神经系统控制。大脑是向神经发送信号的中央信息中心,神经控制并连接运动单元。对于存在的两个不同的运动单元,身体会采用两种不同的神经来控制它们。快速抽搐运动单元由快速抽搐神经控制,而慢速抽搐运动单元由慢速抽搐神经控制。[35]
在实验室中,将运动单元连接到慢肌纤维的神经替换为快肌纤维的神经。慢速拉伸纤维的性能与快速拉伸纤维相同。相反,如果过程相反,那么快拉力光纤也将像慢拉力光纤一样发挥作用。[36]但是,神经不可能从快速运动神经转变为缓慢运动神经,反之亦然。
当人们分析一大群运操作的垂直跳高表现并分析他们的垂直跳高时,就可以证明这一点。 纤维较快的运操作(II型)在运动过程中改变方向的速度较快,例如从左向右,并且他们倾向于较少地屈膝。[37] 这些结果可以通过肌肉活检甚至通过特殊的压板分析来确认。 这并不意味着具有低速弹力纤维的运操作不能跳得更高,但是他们倾向于慢一点并且膝部弯曲程度更大。
另见
Motor learning
Motor skill
Motor coordination
Motor cortex
Multisensory integration
Proprioception
Sensory processing
Two-alternative forced choice
Psychomotor learning
参考
Sibson, Francis (1850). "On The Causes Which Excite And Influence Respiration In Health And Disease". The Transactions of the Provincial Medical and Surgical Association. 5 - New Series: 181–350. In all these instances the act of inspiration is excited through the reflex function of the nervous system -- the sudden impression made on the skin stimulates the extremities of the incident nerves; the stimulus is conveyed by the incident nerves to the spinal nervous centre, and is thence transmitted back over the motor nerves of inspiration. That these respiratory movements are purely excito-motor, and performed without the intervention of sensation, in many of those instances in which the excited movements are most energetic, is proved by the case with which remarkable movements of respiration were occasioned by stimulating the surface in cases of syncope, hysteria, and epilepsy, cases in which sensation was altogether absent, and was only restored after repeatedly stimulating the surface, and so inducing deep reflex inspirations again and again by exciting the incident nerves. [Page 206]
Rosenbaum, David A. (1991). Human 运动控制. San Diego, CA: Academic Press. p. 411. ISBN 978-0-12-597300-7.
Wise, Stephen P.; Shadmehr, Reza (July 10, 2002). "运动控制". Encyclopedia of the Human Brain. Academic Press. pp. 137–157. ISBN 978-0122272103.
Donders, FC. (1969). "On the speed of mental processes" (PDF). Acta Psychol (Amst). 30: 412–31. doi:10.1016/0001-6918(69)90065-1. PMID 5811531.
Hick, W. E. (1952). "On the rate of gain of information". Quarterly Journal of Experimental Psychology. 4 (1): 11–26. doi:10.1080/17470215208416600.
J.A. Adams. A closed loop theory of motor learning. In: Journal of Motor Behavior 3 (1971) p 116. This is also quoted in: Jack A. Adams, Issues for a Closed Loop Theory of Motor Learning. In: George E. Stelmach (Ed.) 运动控制, Issues and Trends. Academic press. New York 1976. p 89
Richard A. Schmidt: 运动控制 and Learning. Human Kinetics Publishers, Champaign, Illinois 1982, p 186
J.A. Adams. A Cllosed Loop Theory of Motor Learning. In: Journal of Motor behavior 3 (1971) p 116. Also quoted in: Jack A. Adams, Issues for a Closed Loop Theory of Motor Learning. In: George E. Stelmach (Ed.) 运动控制, Issues and Trends. Academic press. New York 1976. p 89
Taub, Edward; Steven J. Ellman; A. J. Berman (February 1966). "Deafferentation in Monkeys: Effect on Conditioned Grasp Response". Science. 151 (3710): 593–594. doi:10.1126/science.151.3710.593.
Unni, Midhun P.; Sinha, Aniruddha; Chakravarty, Kingshuk; Chatterjee, Debatri; Das, Abhijit (2017). "Neuromechanical Cost Functionals Governing 运动控制 for Early Screening of Motor Disorders". Frontiers in Bioengineering and Biotechnology. 5: 78. doi:10.3389/fbioe.2017.00078. ISSN 2296-4185. PMC 5733372. PMID 29326926.
Dewhurst DJ (1967). "Neuromuscular control system". IEEE Trans Biomed Eng. 14 (3): 167–71. doi:10.1109/TBME.1967.4502494. PMID 6080533.
Pearson, Keir; Gordon, James (2000), "Spinal reflexes", Principles of Neuroscience, New York, NY: McGraw-Hill, pp. 713–736
Matthews PB (1986). "Observations on the automatic compensation of reflex gain on varying the pre-existing level of motor discharge in man". J Physiol. 374 (1): 73–90. doi:10.1113/jphysiol.1986.sp016066. PMC 1182707. PMID 3746703.
Evarts EV (1973). "Motor cortex reflexes associated with learned movement". Science. 179 (4072): 501–3. doi:10.1126/science.179.4072.501. PMID 4196171.
Latash, Mark L; Scholz, John P; Schner, Gregor (2007). "Toward a new theory of motor synergies". 运动控制. 11 (2): 276–308. doi:10.1123/mcj.11.3.276. PMID 17715460.
Alnajjar, F.; Wojtara, T.; Kimura, H.; Shimoda, S. (2013). "Muscle synergy space: learning model to create an optimal muscle synergy". Frontiers in Computational Neuroscience. 7: 136. doi:10.3389/fncom.2013.00136. PMC 3796759. PMID 24133444.
Latash, Mark (2008). Synergy. Oxford, NY: Oxford University Press. p. 412.
Scholz, John P; Danion, Frederic; Latash, Mark L; Schner, Gregor (2002). "Understanding finger coordination through analysis of the 结构 of force variability". Biological Cybernetics. 86 (1): 29–39. doi:10.1007/s004220100279. PMID 11918210.
Bernstein, Nikolai (1967). The Co-ordination and Regulation of Movement. Long Island City, NY: Permagon Press. p. 196.
Alnajjar, F.; Itkonen, M.; Berenz, V.; Tournier, M.; Nagai, C.; Shimoda, S. (2015). "Sensory synergy as environmental input integration". Frontiers in Neuroscience. 7: 136. doi:10.3389/fnins.2014.00436. PMC 4292368. PMID 25628523.
Henry, Franklin M.; Harrison, John S. (1961). "Refractoriness of Fast Movement". Perceptual and Motor Skills. 13 (3): 351–354. doi:10.2466/pms.1961.13.3.351.
Wadman, W. J.; Denier van der Gon, J. J.; Geuze, R. H.; Mol, C. R. (1979). "Control of Fast Goal-Directed Arm Movements". Journal of Human Movement Studies. 5: 3–17.
Kawato, Mitsuo (1999). "Internal models for 运动控制 and trajectory planning". Current Opinion in Neurobiology. 9 (6): 718–727. doi:10.1016/S0959-4388(99)00028-8. PMID 10607637.
Blakemore, Sarah-Jayne; Daniel Wolpert; Christ Frith (3 August 2000). "Why can't you tickle yourself?". NeuroReport. 11 (11): 11–16. doi:10.1097/00001756-200008030-00002. PMID 10943682.
Shadmehr, Reza; Mussa-Ivaldi, Fernando A. (1994). "Adaptive Re介绍 of Dynamics during Learning of a Motor Task". The Journal of Neuroscience. 14 (5): 3208–3224. doi:10.1523/JNEUROSCI.14-05-03208.1994.
Wolpert, Daniel M.; Ghahramani, Zoubin; Jordan, Michael I. (1995). "An Internal Model for Sensorimotor Integration". Science. 269 (5232): 1880–1882. doi:10.1126/science.7569931. PMID 7569931.
Warren, William H. (2006). "The Dynamics of Perception and Action". Psychological Review. 113 (2): 358–389. CiteSeerX 10.1.1.536.7948. doi:10.1037/0033-295x.113.2.358. PMID 16637765.
Gibson, James J. (1986). The Ecological Approach to Visual Perception. Psychology Press. p. 332. ISBN 978-0898599596.
Michaels, Claire F.; Carello, Claudia (1981). Direct Perception. Englewood Cliffs, NJ: Prentice Hall. p. 200. ISBN 978-0132147910.
Fajen, Brett R.; Warren, William H. (2003). "Behavioral dynamics of steering, obstable avoidance, and route selection". Journal of Experimental Psychology: Human Perception and Performance. 29 (2): 343–362. doi:10.1037/0096-1523.29.2.343.
Fajen, Bret R.; Matthis, Jon S. (2011). "Direct Perception of Action-Scaled Affordances: The Shrinking Gap Problem". Journal of Experimental Psychology: Human Perception and Performance. 37 (5): 1442–1457. doi:10.1037/a0023510. PMC 3140555. PMID 21500936.
Sypert, GW; Munson, JB (1981). "Basis of segmental 运动控制: motoneuron size or motor unit type?". Neurosurgery. 8 (5): 608–621. doi:10.1227/00006123-198105000-00020.
Henneman, Elwood; Somjen, George; Carpenter, David 0. (1965). "Functional Significance of cell size in spinal motor neurons". Journal of Neurophysiology. 28 (3): 560–580. doi:10.1152/jn.1965.28.3.560. PMID 14328454.
Bahler, A. S. (1967). "Series Elastic Component of Mammalian Muscle". American Journal of Physiology. 213 (6): 1560–1564. doi:10.1152/ajplegacy.1967.213.6.1560. PMID 6075758.
Close, R.; Hoh, J. F. (1968). "Effects of nerve cross-union on fast-twitch and slow-graded muscle fibres in the toad". The Journal of Physiology. 198 (1): 103–125. doi:10.1113/jphysiol.1968.sp008596. PMC 1365312. PMID 16992308.
Komi, P. V.; Bosco, C. (1978). "Utilization of stored elastic energy in leg extensor muscles by men and women". Medicine and Science in Sports. 10: 261–265. |

 中国膏药配方配制全书 胡献国,李春日主编P
中国膏药配方配制全书 胡献国,李春日主编P
 推拿按摩的解剖学基础(第4版)美.pdf
推拿按摩的解剖学基础(第4版)美.pdf
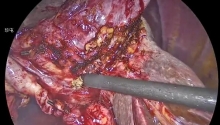
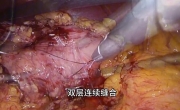 腹腔镜下修复急性器官轴型胃扭转
腹腔镜下修复急性器官轴型胃扭转
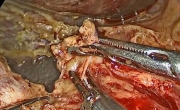 对伴有肝肠系膜干畸形的壶腹腺癌患者行腹腔
对伴有肝肠系膜干畸形的壶腹腺癌患者行腹腔

 发表于 2020-2-2 00:01:28
发表于 2020-2-2 00:01:28
 提升卡
提升卡 置顶卡
置顶卡 沉默卡
沉默卡 喧嚣卡
喧嚣卡 变色卡
变色卡 千斤顶
千斤顶 显身卡
显身卡