马上注册,结交更多好友,享用更多功能,让你轻松玩转社区。
您需要 登录 才可以下载或查看,没有账号?注册
×
神经解剖学是对神经系统的结构和组织的研究。与桡对称的动物的神经系统由分布的细胞网络组成的动物相反,双侧对称的动物具有分离的,明确的神经系统。因此,它们的神经解剖学得到了更好的理解。在脊椎动物中,神经系统被分为大脑和脊髓的内部结构(统称为中枢神经系统,或CNS)以及连接至身体其余部分的神经路径(称为外围神经系统)。 ,或PNS)。对神经系统不同结构和区域的划分对于研究其工作原理至关重要。例如,神经科学家学到的很多东西都来自观察特定大脑区域的损伤或“病变”如何影响行为或其他神经功能。
有关非人类动物神经系统组成的信息,请参见神经系统。有关智人神经系统典型结构的信息,请参见人脑或周围神经系统。本文讨论与神经解剖学研究有关的信息。

神经解剖学是神经系统的解剖学和组织的研究。 图为横截面图,显示了人类大脑的大致解剖结构
内容
1 历史
2 组成
2.1 神经解剖学的方向
3 工具
3.1 细胞染色
3.2 组织化学
3.3 基因编码标记
3.4 非侵入性脑成像
3.5 基于病毒的方法
3.6 染料基方法
3.7 连接组学
3.8 计算神经解剖学
4 模型系统
4.1 秀丽隐杆线虫
4.2 果蝇
5 参考
历史

J. M. Bourgery对大脑,脑干和上脊柱的解剖
关于人类大脑解剖学研究的第一个已知书面记录是古埃及文献,Edwin Smith Papyrus。[1]神经解剖学的下一个重大发展来自希腊人Alcmaeon,他确定大脑而不是心脏统治着身体,并且感觉依赖于大脑。[2]
在Alcmaeon的发现之后,来自世界各地的许多科学家,哲学家和医师继续为神经解剖学的理解做出贡献,尤其是Galen,Herophilus,Rhazes和Erasistratus。亚历山大的赫罗菲罗斯和埃拉西斯特拉特斯可能是最有影响力的希腊神经科学家,他们的研究涉及解剖大脑。[2]此后的数百年中,随着解剖学的文化禁忌,神经科学没有发生重大进展。但是,教皇Sixtus IV通过改变教皇政策并允许人解剖,有效地振兴了神经解剖学研究。这导致了文艺复兴时期的艺术家和科学家对神经解剖学的研究蓬勃发展。[3]
1664年,牛津大学的医师兼教授托马斯·威利斯(Thomas Willis)在发表其著作《脑解剖学》(Cerebri anatome)时创造了“神经病学”一词,这被认为是神经解剖学的基础。[4]随后的三百五十年,产生了大量有关神经系统的文献和研究。
组成
在组织层面,神经系统由神经元,神经胶质细胞和细胞外基质组成。神经元和神经胶质细胞都有许多类型(例如,参见成人体内不同细胞类型列表中的神经系统部分)。神经元是神经系统的信息处理细胞:它们感知作者的环境,通过电信号和称为神经递质的化学物质相互交流,这些物质通常跨突触起作用(两个神经元之间或神经元与肌肉细胞之间的紧密接触;注意突触外作用也是可能的,以及神经递质释放到神经细胞外空间),并产生作者的记忆,思想和动作。胶质细胞维持体内稳态,产生少突胶质细胞,并为大脑的神经元提供支持和保护。一些神经胶质细胞(星形胶质细胞)甚至可以响应刺激而在远距离传播细胞间钙波,并响应钙浓度的变化释放神经胶质递质。大脑中的伤口疤痕主要含有星形胶质细胞。细胞外基质还在分子水平上为大脑细胞提供了支持,使进入和流出血管的物质变质。
在器官一级,神经系统由大脑区域组成,例如哺乳动物中的海马或果蝇的蘑菇体。[5]这些区域通常是模块化的,并在神经系统的一般系统途径中起特定作用。例如,海马对于形成与许多其他大脑区域有关的记忆至关重要。周围神经系统还包含传入或传出神经,它们是源自大脑和脊髓,或源自感觉或运动的周围神经节的纤维束,并反复分支以支配身体的各个部分。神经主要由神经元的轴突或树突构成(如果传出运动纤维,则为轴突;如果传出神经感觉纤维,则为树突),以及包裹它们并分离成神经束的各种膜。
脊椎动物神经系统分为中枢神经系统和外周神经系统。中枢神经系统(CNS)由大脑,视网膜和脊髓组成,而外围神经系统(PNS)由CNS外部的所有神经和神经节(外围神经元的数据包)组成,这些神经和神经节将神经元和神经节连接到神经系统。身体的其余部分。 PNS进一步细分为躯体和自主神经系统。躯体神经系统由“活跃的”神经元和“有效”神经元组成,这些神经元将来自躯体(身体)感觉器官的感觉信息传递给中枢神经系统,而“有效”神经元则将运动指令传送到身体的自愿肌肉。自主神经系统可以在有或没有中枢神经系统控制的情况下工作(这就是为什么被称为“自主”),并且还具有两个细分,称为有交感和副交感神经,这对于将运动命令传递到人体的基本内部器官非常重要。控制心跳,呼吸,消化和流涎等功能。自主神经与体神经不同,仅包含传出纤维。来自内脏的感觉信号通过身体的感觉神经(例如内脏疼痛)或通过某些特定的颅神经(例如化学敏感或机械信号)进入CNS。
神经解剖学的方向

良性家族性大头畸形患者头部的矢状位MRI。
在一般的解剖结构中,尤其是在神经解剖学中,使用了几组地形学术语来表示方向和位置,通常将其称为身体或脑轴(请参见位置解剖学术语)。人们常错误地认为中枢神经系统的轴或多或少是笔直的,但实际上它总是显示出两个腹侧弯曲(子宫颈和头侧弯曲)和背侧弯曲(脑桥弯曲),这都是由于胚胎发生过程中的不同生长所致。在神经解剖学中最常用的术语对是:
背腹:腹侧是指顶板或上侧,以大脑的顶板为代表,腹侧是指板的底侧或下侧,为底板。这些描述符最初用于人体的鼻背和腹膜–背部和腹部。大多数动物的腹部都朝向地面;人类的直立姿势将作者的腹侧向前放置,而背侧则向后移动。头部和大脑的情况是特殊的,因为除非作者假设嘴巴代表腹部的扩展元素,否则腹部无法适当地伸入头部。因此,通常将那些靠近颅骨底部并通过颅骨到达口腔的大脑部分称为腹侧(即如上所定义的在其底部或下侧),而背侧部分则更靠近封闭的颅骨穹顶。提及大脑的顶板和底板不太容易混淆,这也使作者可以密切注意上面提到的轴向弯曲。因此,背侧和腹侧是大脑中的相对术语,其确切含义取决于特定的位置。
鼻状和尾状:鼻状一般是指身体的前部(朝向鼻子或拉丁语的讲台),而尾状是指身体的尾端(朝向尾部;拉丁语为cauda)。大脑的后尾神经尺寸与其长度轴相对应,它的长度轴从脊髓的尾尖横贯所引述的挠曲,一直延伸到视交叉点处的鼻端。在直立的人中,方向性术语“上”和“下”本质上是指该头尾骨尺寸,因为作者的身体和脑轴在直立位置大致垂直。但是,所有的脊椎动物在神经管中都出现非常明显的腹扭结,这种弯曲在成年的中枢神经系统中仍然可以检测到,称为头颅弯曲。后者在前脑(在视交叉处向后终止的轴)与脑干和脊髓(大致垂直的轴,但包括附加轴)之间的过渡处,相对于尾部以180度角弯曲CNS的延髓部。在脑桥和颈椎弯曲处有微小的扭结)当试图描述大脑中的相对位置和切面时,这些轴向尺寸的弯曲变化是有问题的。有大量文献错误地忽略了轴向弯曲,并假设大脑轴相对笔直。
内侧和外侧:内侧是指靠近或相对靠近中线(描述符中值表示恰好位于中线的位置)。横向是相反的(与中线大致分开的位置)。
请注意,此类描述符(背侧/腹侧,鼻侧/尾侧;内侧/外侧)是相对的,而不是绝对的(例如,外侧结构可以说位于内侧,而其他部位甚至位于外侧)。
神经解剖学中取向平面或截面平面的常用术语是“矢状”,“横向”或“冠状”和“轴向”或“水平”。同样在这种情况下,由于轴的位置发生了变化,游泳,爬行或四足动物(俯卧)的情况与人类或其他直立物种的情况有所不同。由于大脑的轴向弯曲,没有一个截面平面能够在选定平面上获得完整的截面系列,因为某些截面不可避免地会在穿过弯曲结构时产生倾斜甚至垂直于其的切口。经验允许辨别导致所需切割的部分。
矢状中平面将身体和大脑分为左右两半。矢状切面通常平行于该中间平面,沿内外侧尺寸移动(请参见上图)。术语“矢状”在词源上是指颅骨左右顶骨之间的正中缝线,在传统上称为矢状缝线,因为它与其他缝线汇合时大致像箭头(矢状;拉丁箭头)。
原则上,与任何细长形式的轴线正交的截面均保持为横向(例如,手指或椎骨的横截面);如果没有长度轴,则无法定义此类截面,或者存在无限可能)。因此,脊椎动物的横断面平行于肋骨,横断面与脊柱正交,该脊柱代表动物和人的体轴。大脑还具有一个固有的纵轴-原始的细长神经管的纵轴-随人的直立姿势在很大程度上垂直,与人体轴相似,除了其鼻端,如上所述。这说明脊髓横切面与作者的肋骨或地面大致平行。但是,这仅适用于脊髓和脑干,因为在早期形态发生过程中,神经轴的前脑末端弯曲成弯曲状,并在其处终止。真实横截面的方向相应地发生变化,不再平行于肋骨和地面,而是垂直于肋骨和地面。对这种形态上的大脑特殊性的认识不足(毫无例外地存在于所有脊椎动物的大脑中)已经引起并且仍然引起对前脑部分的错误思考。认识到延髓横截面的奇异性,传统为它们引入了一个不同的描述词,即冠状横截面。冠状切面将前脑从鼻尖(前)划分为尾(后),形成与局部弯曲轴正交(横向)的序列。由于冠状部分在此处变得与轴向尺寸成水平方向且与轴平行,因此无法将该概念有意义地应用于脑干和脊髓。在任何情况下,“冠状”切面的概念都不如“横向”切面的精确,因为经常使用冠状切面,这些切面并不真正垂直于脑轴的头端。该词在词源上与白杨的冠状缝合相关,并且与戴冠的位置有关(拉丁冠冕指冠)。目前尚不清楚冠的最初含义是什么(可能只是冠冕),不幸的是,这导致了仅定义为冠状的剖面中的歧义。
现代人认为,横跨人头和大脑的冠状面与面部平行(国王的王冠位于其头部的平面与面部并不完全平行,并且该概念向比作者少的前部赋形动物的输出)显然甚至更矛盾,但有一个隐式提及颅骨的冠状缝合线,该缝合线在额骨和颞骨/顶骨之间形成,从而形成了一种大致平行于面部的冠状皱纹。因此,冠状切面实际上仅指有意义的冠状动脉的头部和大脑,而不是指下面的颈部和身体。
根据定义,水平截面与地平线对齐(平行)。在游泳,爬行和四足动物中,体轴本身是水平的,因此,水平部分沿脊髓的长度方向延伸,使腹侧和背侧部分分开。水平截面与横向截面和矢状截面均正交,并且理论上平行于长度轴。由于大脑(前脑)的轴向弯曲,该区域中的真实水平截面与冠状(横向)截面正交(相对于面部的地平线也是如此)。
根据这些考虑,空间的三个方向分别由矢状,横向和水平平面精确表示,而冠状截面可以是横向,倾斜或水平的,具体取决于它们与脑轴及其弯曲的关系。
工具类
神经解剖学的现代发展与用于进行研究的技术直接相关。因此,有必要讨论各种可用的工具。用于研究其他组织的许多组织学技术也可以应用于神经系统。但是,已经开发了一些技术,专门用于神经解剖学的研究。
细胞染色
在生物系统中,染色是一种用于增强显微图像中特定特征对比度的技术。
Nissl染色使用苯胺碱性染料强烈染色粗糙的内质网中的酸性多核糖体,该内质网中神经元含量很高。这使研究人员能够区分神经系统细胞结构各个区域中的不同细胞类型(例如神经元和神经胶质细胞)以及神经元的形状和大小。
经典的高尔基氏染色法使用重铬酸钾和硝酸银选择性地填充铬酸银沉淀物,使一些神经细胞(神经元或神经胶质细胞,但原则上任何细胞都可以类似地反应)。这种所谓的铬酸银浸渍程序可将部分神经元的细胞体和神经突(树突,轴突)染成棕色和黑色,从而使研究人员可以追踪到它们在神经组织切片中最薄的末端分支的路径。透明性的原因是大多数周围细胞都没有染色。现代,高尔基浸渍材料已被用于电子显微镜观察染色过程和细胞体周围的未染色元素,从而增加了分辨力。
组织化学
组织化学利用有关大脑化学成分(主要包括酶)的生化反应特性的知识来应用选择性的反应方法,以可视化它们在大脑中发生的位置以及任何功能或病理变化。这重要地适用于与神经递质产生和代谢有关的分子,但同样适用于许多其他方向,例如化学结构或化学神经解剖学。
免疫细胞化学是组织化学的特例,它使用针对神经系统各种化学表位的选择性抗体来选择性染色特定的细胞类型,轴突束,神经纤维网,神经胶质突或血管,或特定的胞浆内或核内蛋白等免疫遗传分子,例如神经递质。免疫反应的转录因子蛋白揭示了转移蛋白的基因组读数。这极大地提高了研究人员区分神经系统各个区域中不同细胞类型(例如神经元和神经胶质细胞)的能力。
原位杂交使用合成的RNA探针,该探针选择性地附着(杂交)到细胞质中DNA外显子的互补mRNA转录物上,以可视化基因组读数,即根据mRNA而不是蛋白质区分活性基因的表达。这允许组织学上(原位)鉴定参与遗传编码分子生产的细胞,这些分子通常代表分化或功能性状,以及分隔不同脑域或细胞群的分子边界。
基因编码标记
通过在大脑中表达可变数量的红色,绿色和蓝色荧光蛋白,所谓的“彩虹”突变小鼠可以对神经元中许多不同颜色进行组合可视化。这可以标记具有足够独特颜色的神经元,从而通常可以通过荧光显微镜将它们与邻居区分开来,从而使研究人员能够绘制神经元之间的局部连接或相互排列(平铺)。
光遗传学使用封闭标记的转基因组成型和位点特异性表达(通常在小鼠中),可以通过光束照射选择性激活。这使研究人员能够以非常有区别的方式研究神经系统中的轴突连接性。
非侵入性脑成像
磁共振成像已广泛用于研究健康人的大脑结构和非侵入性功能。一个重要的例子是扩散张量成像,其依靠水在组织中的受限扩散来产生轴突图像。特别是,水沿着与轴突对齐的方向移动得更快,从而可以推断其结构。
基于病毒的方法
某些病毒可以在脑细胞中复制并发生突触。因此,经过修饰以表达标记物(例如荧光蛋白)的病毒可用于追踪多个突触的大脑区域之间的连通性。[6]复制和传播跨神经元/突触的两种示踪病毒是单纯疱疹病毒1型(HSV)[7]和弹状病毒[8]。单纯疱疹病毒被用来追踪大脑和胃之间的联系,以检查涉及内脏感觉加工的大脑区域。[9]另一项研究将单纯疱疹病毒注射入眼睛,从而使从视网膜到视觉系统的光路可视化。[10]从突触复制到体细胞的示踪病毒的一个例子是伪狂犬病病毒。[11]通过使用带有不同荧光报告基因的伪狂犬病病毒,双重感染模型可以解析复杂的突触结构。[12]
染料基方法
轴突运输方法使用多种染料(或多或少地被神经元或其过程吸收)(辣根过氧化物酶变体,荧光或放射性标记,凝集素,右旋糖酐)。这些分子被选择性地顺行地运输(从躯体到轴突末端)或逆行地运输(从轴突终端到躯体),从而提供了大脑中主要和间接连接的证据。这些“生理的”方法(由于使用了活的,未损伤的细胞的特性)可以与其他方法结合使用,并且已基本取代了先前研究损伤神经元或轴突变性的早期方法。详细的突触连接可以通过相关电子显微镜确定。
连接组学
主条目:连接组学
串行截面电子显微镜已被广泛开发用于研究神经系统。例如,串行块面扫描电子显微镜的首次应用是在啮齿动物的皮质组织上。[13]用这种高通量方法产生的数据进行电路重构具有挑战性,并且开发了公民科学游戏EyeWire来辅助该领域的研究。
计算神经解剖学
主要文章:计算神经科学
是一个利用各种成像方式和计算技术来对正常和临床人群中神经解剖结构的时空动态进行建模和量化的领域。
模型系统
除人脑外,还有许多其他动物的大脑和神经系统已作为模型系统得到广泛研究,包括小鼠,斑马鱼,[14]果蝇[15]和一种名为秀丽隐杆线虫的of虫。作为模型系统,每种方法都有其自身的优点和缺点。例如,秀丽隐杆线虫的神经系统从一个蠕虫到另一个蠕虫的刻板印象。这使得研究人员可以使用电子显微镜来绘制该物种中所有大约300个神经元的路径和连接。对果蝇进行了广泛的研究,部分原因是它的遗传学得到了很好的理解并且易于操纵。使用鼠标是因为,作为哺乳动物,它的大脑在结构上与作者的大脑更相似(例如,它具有六层皮质,但是其基因很容易被修饰,并且其繁殖周期相对较快)。
秀丽隐杆线虫
杆状体包含一个消化系统,该消化系统从一端的嘴巴延伸到另一端的肛门。沿着消化系统的是一条神经索,末端是大脑,靠近嘴。

普通双语动物的神经系统,呈神经节的形式,具有节段性扩大,并且在前部有“大脑”
在某些物种中,大脑小而简单,例如线虫蠕虫,其身体计划非常简单:从嘴到肛门有一个带有中空肠腔的管,以及一个膨大的神经索(神经节)。对于每个身体部分,在前面都有一个特别大的神经节,称为大脑。由于线虫秀丽隐杆线虫在遗传学中的重要性,因此对其进行了研究。[16]在1970年代初,悉尼布伦纳(Sydney Brenner)选择它作为模型系统来研究基因控制发育的方式,包括神经元发育。使用这种蠕虫的一个优势是,雌雄同体的神经系统恰好包含302个神经元,始终位于相同的位置,从而在每种蠕虫中都具有相同的突触连接。[17]布伦纳(Brenner)的团队将蠕虫切成数千个超薄切片,并在电子显微镜下对每个切片进行拍照,然后在视觉上匹配每个切片的纤维,以绘制出整个身体的每个神经元和突触,从而形成一个完整的线虫连接体。[18] ]没有任何其他生物能够达到如此详细的水平,并且该信息已被用于进行大量的研究,而没有这些研究就无法进行。[19]
果蝇
更多信息:果蝇连接组
果蝇(Drosophila melanogaster)是一种流行的实验动物,因为它很容易从野外大规模培养,产生时间短,并且容易获得突变动物。
节肢动物的中央大脑具有三个部分,每只眼睛后面都有大的光波瓣,用于视觉处理。果蝇的大脑包含数百万个突触,而人脑中至少有1,000亿个突触。果蝇大脑的大约三分之二专用于视觉处理。
托马斯·亨特·摩根(Thomas Hunt Morgan)于1906年开始与果蝇合作,这项工作为他赢得了1933年诺贝尔医学奖,因为他将染色体鉴定为基因的遗传载体。由于可用于研究果蝇遗传学的工具众多,它们已成为研究基因在神经系统中作用的自然学科。[20]该基因组已测序并于2000年发表。大约75%的已知人类疾病基因在果蝇的基因组中具有可识别的匹配。果蝇被用作几种人类神经系统疾病的遗传模型,包括帕金森氏病,亨廷顿氏病,脊髓小脑共济失调和阿尔茨海默氏病等神经退行性疾病。尽管昆虫和哺乳动物之间的进化距离很大,但果蝇神经遗传学的许多基本方面已证明与人类有关。例如,通过检查果蝇突变体,发现其日常活动周期被破坏,从而鉴定出第一个生物钟基因。[21]
另见
Connectogram
Outline of the human brain
Outline of brain mapping
List of regions in the human brain
Medical image computing
Neurology
Neuroscience
Computational anatomy
参考
Atta, H. M. (1999). "Edwin Smith Surgical Papyrus: The Oldest Known Surgical Treatise". American Surgeon. 65 (12): 1190–1192.
Rose, F (2009). "Cerebral Localization in Antiquity". Journal of the History of the Neurosciences. 18 (3): 239–247. doi:10.1080/09647040802025052. PMID 20183203.
Ginn, S. R.; Lorusso, L. (2008). "Brain, Mind, and Body: Interactions with Art in Renaissance Italy". Journal of the History of the Neurosciences. 17 (3): 295–313. doi:10.1080/09647040701575900.
Neher, A (2009). "Christopher Wren, Thomas Willis and the Depiction of the Brain and Nerves". Journal of Medical Humanities. 30 (3): 191–200. doi:10.1007/s10912-009-9085-5. PMID 19633935.
Mushroom Bodies of the Fruit Fly Archived 2012-07-16 at Archive.today
Ginger, M.; Haberl, M.; Conzelmann, K.-K.; Schwarz, M.; Frick, A. (2013). "Revealing the secrets of neuronal circuits with recombinant rabies virus technology". Front. Neural Circuits. 7: 2. doi:10.3389/fncir.2013.00002. PMC 3553424. PMID 23355811.
McGovern, AE; Davis-Poynter, N; Rakoczy, J; Phipps, S; Simmons, DG; Mazzone, SB (2012). "Anterograde neuronal circuit tracing using a genetically modified herpes simplex virus expressing EGFP". J Neurosci 方法. 209 (1): 158–67. doi:10.1016/j.jneumeth.2012.05.035. PMID 22687938.
Kuypers HG, Ugolini G (February 1990). "Viruses as transneuronal tracers". Trends in Neurosciences. 13 (2): 71–5. doi:10.1016/0166-2236(90)90071-H. PMID 1690933.
Rinaman L, Schwartz G (March 2004). "Anterograde transneuronal viral tracing of central viscerosensory pathways in rats". The Journal of Neuroscience. 24 (11): 2782–6. doi:10.1523/JNEUROSCI.5329-03.2004. PMID 15028771.
Norgren RB, McLean JH, Bubel HC, Wander A, Bernstein DI, Lehman MN (March 1992). "Anterograde transport of HSV-1 and HSV-2 in the visual system". Brain Research Bulletin. 28 (3): 393–9. doi:10.1016/0361-9230(92)90038-Y. PMID 1317240.
Card, J. P. (2001). "Pseudorabies virus neuroinvasiveness: A window into the functional organization of the brain". Advances in Virus Research. 56: 39–71. doi:10.1016/S0065-3527(01)56004-2. ISBN 9780120398560.
Card, J. P. (2011). "A Dual Infection Pseudorabies Virus Conditional Reporter Approach to Identify Projections to Collateralized Neurons in Complex Neural Circuits". PLoS ONE. 6 (6): e21141. doi:10.1371/journal.pone.0021141. PMC 3116869. PMID 21698154.
Denk, W; Horstmann, H (2004). "Serial Block-Face Scanning Electron Microscopy to Reconstruct Three-Dimensional Tissue Nano结构". PLoS Biology. 2 (11): e329. doi:10.1371/journal.pbio.0020329. PMC 524270. PMID 15514700.
Wullimann, Mario F.; Rupp, Barbar; Reichert, Heinrich (1996). Neuroanatomy of the zebrafish brain: a topological atlas. ISBN 3-7643-5120-9. Archived from the original on 2013-06-15. Retrieved 2016-10-16.
"Atlas of the Drosophila Brain". Archived from the original on 2011-07-16. Retrieved 2011-03-24.
"WormBook: The online review of C. elegans biology". Archived from the original on 2011-10-11. Retrieved 2011-10-14.
Hobert, Oliver (2005). The C. elegans Research Community (ed.). "Specification of the nervous system". WormBook: 1–19. doi:10.1895/wormbook.1.12.1. PMC 4781215. PMID 18050401. Archived from the original on 2011-07-17. Retrieved 2011-11-05.
White, JG; Southgate, E; Thomson, JN; Brenner, S (1986). "The 结构 of the Nervous System of the Nematode Caenorhabditis elegans". Philosophical Transactions of the Royal Society B. 314 (1165): 1–340. doi:10.1098/rstb.1986.0056. PMID 22462104.
Hodgkin, J (2001). "Caenorhabditis elegans". In Brenner S, Miller JH (eds.). Encyclopedia of Genetics. Elsevier. pp. 251–256. ISBN 978-0-12-227080-2.
"Flybrain: An online atlas and database of the drosophila nervous system". Archived from the original on 2016-05-16. Retrieved 2011-10-14.
Konopka, RJ; Benzer, S (1971). "Clock Mutants of Drosophila melanogaster". Proc. Natl. Acad. Sci. U.S.A. 68 (9): 2112–6. doi:10.1073/pnas.68.9.2112. PMC 389363. PMID 5002428. |

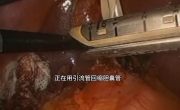 腹腔镜下胆总管囊肿切除术及高级胆道重建术
腹腔镜下胆总管囊肿切除术及高级胆道重建术
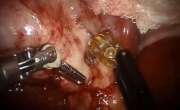 机器人辅助腹腔镜胆囊次全切除术和胆总管探
机器人辅助腹腔镜胆囊次全切除术和胆总管探
 困难腹腔镜胆囊切除术中的吲哚菁绿荧光胆管
困难腹腔镜胆囊切除术中的吲哚菁绿荧光胆管
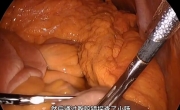 腹腔镜治疗85岁女性胆石性肠梗阻
腹腔镜治疗85岁女性胆石性肠梗阻

 发表于 2019-12-31 00:00:36
发表于 2019-12-31 00:00:36
 提升卡
提升卡 置顶卡
置顶卡 沉默卡
沉默卡 喧嚣卡
喧嚣卡 变色卡
变色卡 千斤顶
千斤顶 显身卡
显身卡