马上注册,结交更多好友,享用更多功能,让你轻松玩转社区。
您需要 登录 才可以下载或查看,没有账号?注册
×
分子神经科学是神经科学的一个分支,它观察应用于动物神经系统的分子生物学概念。 该主题的范围包括诸如分子神经解剖学,神经系统中分子信号传导的机制,遗传学和表观遗传学对神经元发育的影响以及神经可塑性和神经退行性疾病的分子基础等主题。[1] 与分子生物学一样,分子神经科学是一个相对较新的领域,它具有很大的动态性。
内容
1 定位神经递质
2 电压门控离子通道
2.1 钠离子通道
2.2 钾离子通道
2.3 钙离子通道
3 受体
3.1 离子受体
3.1.1 GABA受体
3.1.2 谷氨酸受体
3.1.3 烟碱型ACh受体
3.2 代谢型受体
3.2.1 G蛋白连锁受体
4 神经递质释放
4.1 神经递质的释放依赖钙
5 神经元基因表达
5.1 性别差异
5.2 大脑的表观遗传学
6 神经退行性疾病的分子机制
6.1 兴奋性毒性和谷氨酸受体
6.2 阿尔茨海默氏病
6.3 帕金森氏病
6.4 亨廷顿舞蹈病
7 参考
定位神经递质
在分子生物学中,神经元之间的交流通常通过跨细胞间隙的化学传递(称为突触)发生。所传播的化学物质称为神经递质,可调节重要的人体机能。[2]通过标记技术可以解剖定位神经递质。通过用甲醛固定神经组织切片,可以化学鉴定某些神经递质,例如儿茶酚胺。当暴露于紫外线下时,这会引起甲醛诱导的荧光。使用该技术可在线虫秀丽隐杆线虫中鉴定出多巴胺,一种儿茶酚胺。[3]免疫细胞化学涉及产生针对靶向化学或生物实体的抗体,还包括其他一些令人感兴趣的技术。靶向的神经递质可以由一抗和二抗进行放射性标记,从而通过放射自显影来识别神经递质。可以在酶联免疫细胞化学或酶联免疫吸附测定(ELISA)中观察到神经递质的存在(尽管不一定是位置),其中酶联测定中的底物结合可以诱导沉淀物,荧光团或化学发光。如果无法通过组织化学方法鉴定神经递质,一种替代方法是通过其神经摄取机制对其进行定位。[1]
电压门控离子通道

真核电压门控钾离子通道的结构
活生物体中的可激发细胞具有电压门控离子通道。这些可以在神经元的整个神经系统中观察到。 1950年代,A.L。Hodgkin和A.F. Huxley研究了鱿鱼属Loligo的巨大轴突后,第一个被表征的离子通道是钠离子和钾离子通道。他们的研究证明了细胞膜的选择性渗透性,取决于生理条件,以及这些渗透性产生动作电位所产生的电效应。[4]
钠离子通道
钠通道是1984年由Shosaku Numa从鳗鱼Electrophorus electrous分离的第一个电压门控离子通道。河豚毒素河豚毒素(TTX)是一种钠通道阻滞剂,通过使用色谱法进行化学分离,将其与钠通道蛋白结合来分离钠通道蛋白。通过Edman降解分析该蛋白的氨基酸序列,然后用于构建可用于克隆通道蛋白的cDNA文库。克隆通道本身可用于诸如识别其他动物的相同通道等应用。[1]钠离子通道与钾离子通道在梯度电位和动作电位的形成过程中协同作用。钠通道允许Na +离子流入神经元,从而导致神经元的静息膜电位发生去极化,从而导致分级的电位或动作电位,具体取决于去极化的程度。[5]
钾离子通道
钾通道以多种形式存在,存在于大多数真核细胞中,并且通常倾向于将细胞膜稳定在钾平衡电位下。与钠离子一样,梯度电位和动作电位也取决于钾离子通道。尽管Na +离子流入神经元会引起细胞去极化,但K +离子流出神经元会导致细胞重新极化至静止的膜电位。钾离子通道的激活本身取决于动作电位期间Na +流入引起的去极化。[1]与钠通道一样,钾通道也有自己的毒素,这些毒素会阻断通道蛋白的作用。这种毒素的一个例子是大阳离子四乙铵(TEA),但是值得注意的是,鉴于物种间不同的通道类型,该毒素对所有钾通道的作用机理不同。钾通道的存在首先在果蝇的果蝇突变体中发现,果蝇由于细胞复极化的问题导致神经元和肌肉电生理异常,在麻醉后无法控制地摇动。钾通道首先是通过操纵(果蝇)的分子遗传学而不是进行通道蛋白纯化来鉴定的,因为在发现时尚不存在钾通道(例如TEA)的已知高亲和力配体。[1] [6]
钙离子通道
钙通道对于某些细胞信号级联以及轴突末端的神经递质释放非常重要。在可兴奋细胞中发现了多种不同类型的钙离子通道。与钠离子通道一样,已经通过色谱纯化技术分离并克隆了钙离子通道。值得注意的是,与神经递质的释放一样,钙通道可以与细胞内蛋白质相互作用,并且在信号传导中起着重要作用,尤其是在肌肉细胞的肌浆网等位置。[1]
受体
各种类型的受体可用于细胞信号传导和通讯,并可包括离子型和代谢型受体。 这些细胞表面受体的类型通过作用机理和作用持续时间来区分,其中离子性受体与快速信号传输有关,而代谢型受体与缓慢的信号传输有关。 代谢型受体恰好覆盖了具有明显不同信号级联的多种细胞表面受体。[1] [5]
离子受体

Ca2 +离子流情况下离子型受体的原型描述
离子型受体,也称为配体门控离子通道,是速效受体,通过离子通道与配体结合而介导神经和生理功能。烟碱,GABA和谷氨酸受体是受配体门控离子通道流动调节的一些细胞表面受体。 GABA是大脑的主要抑制性神经递质,而谷氨酸是大脑的主要兴奋性神经递质。[1]
GABA受体
已知GABAA和GABAC受体是离子性的,而GABAB受体是亲代谢的。 GABAA受体介导中枢神经系统(CNS)的快速抑制反应,并存在于神经元,神经胶质细胞和肾上腺髓质细胞上。它负责诱导Cl离子流入细胞,从而降低了在梯度电位或动作电位到达时发生膜去极化的可能性。 GABA受体也可以与非内源性配体相互作用以影响活性。例如,化合物安定(以安腾(安定)出售)是一种变构激动剂,可增加受体对GABA的亲和力。 GABA结合增加导致的生理抑制作用增强,使安定成为有用的镇定剂或抗惊厥药(抗癫痫药)。另一方面,GABA受体也可以通过减少Cl细胞的流入而成为靶点,并具有诸如微毒素之类的惊厥作用。该化合物的拮抗作用机理并不直接作用于GABA受体,而是存在其他能够变构失活的化合物,包括T-丁基双环佛硫代酸酯(TBPS)和戊四氮(PZT)。与GABAA相比,GABAC受体对GABA的亲和力更高,它们的活性可能更持久,并且其响应可能由较低的GABA浓度产生。[1]
谷氨酸受体
离子型谷氨酸受体可以包括NMDA,AMPA和海藻酸酯受体。这些受体以促进谷氨酸活性的激动剂命名。 NMDA受体的兴奋机制会影响学习和记忆中的神经元可塑性,以及中风和癫痫等神经病理学。 NDMA受体就像离子型GABA受体一样具有多个结合位点,并且会受到诸如甘氨酸神经递质或苯环利定(PCP)等共激动剂的影响。 NMDA受体通过Ca2 +离子流过电流,并可能被细胞外Mg2 +离子阻断,具体取决于电压和膜电位。 NMDA受体产生的兴奋性突触后电位(EPSP)激活基于Ca2 +的信号级联反应(例如神经递质释放),从而增加了Ca2 +的内流。与其他离子型谷氨酸受体相比,AMPA产生更短,更大的兴奋性突触后电流。[5]
烟碱乙酰胆碱受体
烟碱受体与乙酰胆碱(ACh)神经递质结合,产生产生兴奋性突触后反应的非选择性阳离子通道。受体活性可能会受到尼古丁消耗的影响,产生高高兴兴,放松和不可避免的成瘾感。[5]
代谢型受体

G蛋白连锁受体信号转导
代谢型受体是突触后细胞中的慢反应受体。通常,这些缓慢的反应的特征是生物化学中细胞内更精细的变化。代谢型受体对神经递质摄取的反应会导致细胞内酶的激活和涉及第二信使的级联反应,与G蛋白连接的受体一样。各种代谢型受体可包括某些谷氨酸受体,毒蕈碱型ACh受体,GABAB受体和酪氨酸激酶。
G蛋白连锁受体
与G蛋白连接的信号级联反应可以显著放大特定神经递质的信号,从而在细胞中产生数百至数千个第二信使。 G蛋白连接的受体引起信号级联的作用机理如下:
神经递质与受体结合
受体发生构象变化以允许G蛋白复合物结合
G蛋白复合物与受体结合后,GDP与GTP交换
G蛋白复合物的α亚基与GTP结合并分离以与靶蛋白(例如腺苷酸环化酶)结合
与靶蛋白的结合会增加或降低第二信使(例如环AMP)的产生速率
GTPase水解α-亚基,使其与GDP结合,α-亚基返回无活性的G蛋白复合体
神经递质释放

发生神经递质释放和摄取的突触的结构
神经递质从一个神经元的轴突末端到整个神经突触的树突之间以离散量释放,称为量子。这些量子已通过电子显微镜鉴定为突触小泡。两种类型的囊泡分别是直径约40-60nm的小突触囊泡(SSV)和直径约120-200nm的大密芯囊泡(LDCV)电子密集囊泡[1]。前者源自内体,并含有神经递质,例如乙酰胆碱,谷氨酸,GABA和甘氨酸。后者来自高尔基体,并容纳较大的神经递质,例如儿茶酚胺和其他肽类神经递质。[7]神经递质从轴突末端释放并在以下过程中与突触后树突结合:[5]
从细胞骨架操作/招募突触小泡
囊泡(结合)对突触前膜的对接
通过ATP引发囊泡(相对较慢的步骤)
初免囊泡与突触前膜的融合和所容纳神经递质的胞吐作用
突触后细胞受体中神经递质的摄取
取决于神经递质是兴奋性的还是抑制性的,在突触后细胞中动作电位的起始或抑制(兴奋将导致突触后膜去极化)
神经递质的释放依赖钙
神经递质的释放取决于外部Ca2 +离子的供应,这些Ca2 +离子通过电压门控钙通道进入轴突末端。与末端膜的囊泡融合和神经递质的释放是由传入动作电位引起的Ca2 +梯度的产生引起的。 Ca 2+离子导致从储备池中新合成的囊泡操作,从而进行这种膜融合。这种作用机理是在鱿鱼巨型轴突中发现的。[8]降低细胞内Ca2 +离子可直接抑制神经递质的释放[1]。在神经递质释放发生后,囊泡膜被回收到其生产来源。钙离子通道可根据入射位置而变化。例如,轴突末端的通道不同于细胞体的典型钙通道(无论是神经通道还是非神经通道)。即使在轴突末端,钙离子通道的类型也会发生变化,就像位于神经肌肉接头处的P型钙通道一样。[1]
神经元基因表达
另见:计算神经遗传学建模
性别差异
性别决定上的差异由性别染色体控制。性激素释放对大脑的性二态性(性特征的表型分化)有重要影响。最近的研究似乎表明,调节这些二态性对理解正常和异常的脑功能有影响。性二态性可能受到基于性别的大脑基因表达的影响,该性别基因因物种而异。
动物模型(例如啮齿动物,黑腹果蝇和秀丽隐杆线虫)已用于观察大脑中与动物产生激素的性腺相对的性别偏见的起源和/或程度。对于啮齿类动物,对性染色体进行基因操作的研究导致对一种性别的影响与另一种性别的影响完全相反。例如,敲除特定基因只会在男性中引起焦虑样的影响。通过对D. menlanogaster的研究,发现即使在去除性腺后,仍会出现较大的大脑性别表达偏见,这表明性别偏见在某些方面可能与激素控制无关。[9]
观察具有性别偏见的基因具有观察脑生理的临床意义和潜在的相关(无论是直接还是间接)神经系统疾病的潜力。在发育中带有性别偏见的疾病包括亨廷顿氏病,脑缺血和阿尔茨海默氏病。[9]
大脑的表观遗传学
基因表达的变化和改变会在细胞和分子水平上影响许多大脑功能,而不会改变生物体中DNA的序列。这就是所谓的表观遗传调控。表观遗传机制的例子包括组蛋白修饰和DNA甲基化。已经发现这种变化对脑疾病,精神疾病和成瘾的发生有很大影响。[10]已证明表观遗传控制与早期发育中的高水平可塑性有关,从而确定了其在生物关键时期的重要性。[11]表观遗传变化如何影响人脑的示例如下:
脑海马中rRNA基因的甲基化水平较高,结果导致蛋白质产量降低,因此海马功能受限会导致学习和记忆障碍以及自杀倾向。[12]
在一项比较健康人与精神病患者之间遗传差异的研究中,发现了60种与脑细胞信号传导相关的表观遗传标记。[12]
诸如虐待儿童之类的环境因素似乎导致在自杀受害者中未发现的糖皮质激素受体上表达表观遗传标签(与压力反应相关)。[12]这是依赖于经验的可塑性的一个例子。
个体的环境富集与海马基因组蛋白乙酰化增加有关,从而改善了记忆巩固(特别是空间记忆)。[11]
神经退行性疾病的分子机制
兴奋性毒性和谷氨酸受体
兴奋毒性是谷氨酸受体被不适当激活的现象。这可能是由于长时间的兴奋性突触传递所致,其中高水平的谷氨酸神经递质导致突触后神经元的过度活化,从而导致突触后神经元的死亡。在脑损伤后(例如由于缺血),已经发现兴奋性毒性是神经元损伤的重要原因。如果在缺血期间由于谷氨酸和天冬氨酸含量增加而导致大脑血液流量减少后突然灌注血液会导致过度的突触活动,这是可以理解的。[5] [13]
阿尔茨海默氏病
阿尔茨海默氏病是最常见的神经退行性疾病,并且是老年人痴呆症的最常见形式。该疾病的特征在于进行性记忆的丧失和各种认知功能。据推测,淀粉样蛋白-β肽(40-42个氨基酸残基)在大脑中的沉积是阿尔茨海默氏病发病所必需的。据称积累会阻止海马的长期增强。淀粉样β寡聚体的受体也可能是a病毒蛋白。[14]
帕金森氏病
帕金森氏病是仅次于阿尔茨海默氏病的第二大最常见的神经退行性疾病。这是由于人脑黑质中多巴胺能神经元的丧失而引起的运动不足的运动性基底神经节疾病。因此,基底神经节的抑制性流出并没有减少,因此由丘脑介导的上运动神经元没有及时激活。具体症状包括僵硬,姿势问题,动作缓慢和震颤。阻断GABA受体从中棘神经元到网状细胞的输入,会导致上运动神经元的抑制作用类似于帕金森氏病[5]。
亨廷顿氏病
亨廷顿舞蹈病是一种运动过度运动引起的基底神经节疾病,是由于缺乏来自基底神经节的中棘状神经元的正常抑制性输入所致。这带来了与帕金森氏病相关的相反作用,包括上运动神经元的不适当激活。与观察到的与帕金森氏病有关的GABA能机制一样,向黑质部网状结构中注射的GABA激动剂减少了对上运动神经元的抑制,导致弹道非自愿运动,类似于亨廷顿舞蹈病的症状。[5]
参考
Longstaff, Alan; Revest, Patricia (1998). Molecular Neuroscience. Garland Science. ISBN 978-1859962503.
"What are Neurotransmitters?". Retrieved 1 November 2013.
Riddle, Donald (1998). C. Elegans II. New York: Cold Spring Harbor Laboratory Press. ISBN 978-0879695323.
Hodgkin, Allan L.; Andrew F. Huxley (1952). "The dual effect of membrane potential on sodium conductance in the giant axon of Loligo" (PDF). The Journal of Physiology. 116 (4): 497–506. doi:10.1113/jphysiol.1952.sp004719. PMC 1392212. PMID 14946715.
Purves, Dale (2012). Neuroscience (5th ed.). Massachusetts, USA: Sinauer Associates, Inc. p. 80. ISBN 978-0-87893-695-3.
Kamb, Alexander; Linda E. Iverson; Mark A. Tanouye (31 July 1987). "Molecular characterization of Shaker, a Drosophila gene that encodes a potassium channel". Cell. 50 (3): 405–413. doi:10.1016/0092-8674(87)90494-6. PMID 2440582.
Davies, R. Wayne; Brian J. Morris (1997). Molecular Biology of the Neuron. Oxford, UK: BIOS Scientific Publishers Ltd. ISBN 978-1859962404.
Dipolo, R.; C. Caputo; F. Bezanilla (March 1983). "Voltage-dependent calcium channel in the squid axon". Proc Natl Acad Sci U S A. 80 (6): 1743–1745. Bibcode:1983PNAS...80.1743D. doi:10.1073/pnas.80.6.1743. PMC 393680. PMID 6300873.
Jazin, E.; Cahill, L. (January 2010). "Sex differences in molecular neuroscience: from fruit flies to humans". Nature Reviews Neuroscience. 11 (1): 9–17. doi:10.1038/nrn2754. PMID 20019686.
"Epigenetics and the Human Brain". Genetics Science and Learning Center at The University of Utah. Retrieved 10 November 2013.
Fagiolini, Michela; Catherin L. Jensen; Frances A. Champagne (2009). "Epigenetic Influences on brain development and plasticity" (PDF). Current Opinion in Neurobiology. 19 (2): 1–6. doi:10.1016/j.conb.2009.05.009. PMC 2745597. PMID 19545993. Retrieved 14 November 2013.
Tsankova, Nadia; Renthal, William; Kumar, Arvind; Nestler, Eric J. (2007). "Epigenetic Regulation in Psychiatric Disorders". Nature Reviews Neuroscience. 8 (5): 355–367. doi:10.1038/nrn2132. PMID 17453016.
Lau, A.; M. Tymianski (2010). "Glutamate receptors, neurotoxicity and neurodegeneration". Pflügers Arch. 460 (2): 525–542. doi:10.1007/s00424-010-0809-1. PMID 20229265.
Laren, Juha; David A. Gimbel; Haakon B. Nygaard; John W. Gilbert (February 2009). "Cellular prion protein mediates impairment of synaptic plasticity by amyloid-β oligomers". Nature. 457 (7233): 1128–1132. Bibcode:2009Natur.457.1128L. doi:10.1038/nature07761. PMC 2748841. PMID 19242475. |

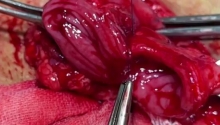
 因缺血而行小肠分段切除及一期吻合术
因缺血而行小肠分段切除及一期吻合术
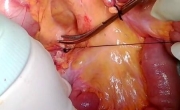
 肌肉活检 _ 如何进行肌肉活检
肌肉活检 _ 如何进行肌肉活检
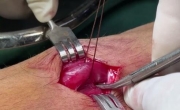
 尿道下裂修复后包皮瘘和龟头隐裂
尿道下裂修复后包皮瘘和龟头隐裂
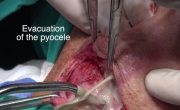 通过外部方法对喉囊肿进行有袋化或去顶
通过外部方法对喉囊肿进行有袋化或去顶

 发表于 2019-12-29 00:00:26
发表于 2019-12-29 00:00:26
 提升卡
提升卡 置顶卡
置顶卡 沉默卡
沉默卡 喧嚣卡
喧嚣卡 变色卡
变色卡 千斤顶
千斤顶 显身卡
显身卡