马上注册,结交更多好友,享用更多功能,让你轻松玩转社区。
您需要 登录 才可以下载或查看,没有账号?注册
×
小胶质细胞是遍布大脑和脊髓的一种神经胶质细胞(胶质细胞)。[1]小胶质细胞占脑内发现的所有细胞的10–15%。[2]作为常驻巨噬细胞,它们是中枢神经系统(CNS)主动免疫防御的第一种也是主要形式。[3]小胶质细胞(以及包括星形胶质细胞在内的其他神经胶质细胞)分布在整个CNS的较大的非重叠区域。[4] [5]小胶质细胞是整个大脑维护的关键细胞,它们不断清除中枢神经系统的斑块,受损或不必要的神经元和突触以及感染因子。[6]由于这些过程必须有效防止潜在的致命伤害,因此小胶质细胞对中枢神经系统的微小病理变化极为敏感。[7]这种敏感性部分是由于独特的钾通道的存在而实现的,这些通道可对细胞外钾的微小变化做出反应。[6]
构成中枢神经系统的大脑和脊髓,由于一系列称为血脑屏障或BBB的内皮细胞,通常不会被人体循环系统中的病原体直接进入。 BBB可防止大多数感染到达脆弱的神经组织。如果将传染原直接引入大脑或穿过血脑屏障,则小胶质细胞必须迅速反应以减少炎症并破坏传染原,然后才能损害敏感的神经组织。由于身体其余部位缺乏抗体(很少有抗体足够小,无法穿过血脑屏障),小胶质细胞必须能够识别异物,吞下它们,并充当激活T细胞的抗原呈递细胞。

创伤性脑损伤前大鼠皮质的小胶质细胞处于静止状态(HRP凝集素染色)

小胶质细胞/巨噬细胞–大鼠脑部创伤后皮层的活化形式(HRP凝集素染色)
内容
1 类型
2 性别遗传学
2.1 分枝
2.2 反应式(已激活)
2.2.1 非吞噬
2.2.2 吞噬
2.3 变形虫
2.4 发光单元
2.5 血管周围
2.6 脉管
3 功能
3.1 清除
3.2 吞噬作用
3.3 细胞外信号
3.3.1 抗原介绍
3.3.2 细胞毒性
3.3.3 突触剥离
3.3.4 促进修复
4 发展
4.1 老化
5 临床意义
6 历史
7 参考
类型

大鼠小胶质细胞在组织培养中以绿色生长,神经纤维过程以红色显示。

大鼠小脑分子层中的小胶质细胞为红色,用针对IBA1 / AIF1的抗体染色。 Bergmann胶质细胞过程以绿色显示,DNA以蓝色显示。
小胶质细胞具有极强的可塑性,并且会根据位置和系统需求而发生各种结构变化。要实现小胶质细胞执行的各种功能,就需要这种可塑性。转化的能力将小胶质细胞与巨噬细胞区分开来,巨噬细胞必须定期更换,并赋予它们在极短的时间内保护中枢神经系统的能力,而不会引起免疫干扰。[6]小胶质细胞根据当地条件和他们检测到的化学信号采用特定的形式或表型。[8]
感性遗传学
小胶质细胞的感觉是一个相对较新的生物学概念,似乎在神经发育和神经变性中起着重要作用。感觉性是指用于感测配体和微生物的蛋白质转录物的独特分组。换句话说,感官代表用于感测体内分子的蛋白质所需的基因。可以使用多种方法来分析感官,包括qPCR,RNA-seq,微阵列分析和直接RNA测序。浆膜上的受体和跨膜蛋白的感官编码中包含的基因与神经元相比在小胶质细胞中表达更高。它不包括特定于膜结合细胞器的分泌蛋白或跨膜蛋白,例如细胞核,线粒体和内质网。[9]多个已鉴定的感官基因编码模式识别受体,但是,包含的基因种类繁多。小胶质细胞与其他巨噬细胞具有相似的感觉,但是它们包含22个独特的基因,其中16个用于与内源性配体相互作用。这些差异创建了一个独特的小胶质生物标记,其中包括40多个基因,包括P2ry12和HEXB。 DAP12似乎在感觉蛋白相互作用中起重要作用,充当信号转导接头和调节蛋白。[9]
感官内部基因的调节必须能够改变,以应对潜在的伤害。小胶质细胞可以承担神经保护或神经毒性的作用,以应对这些危险。[10]由于这些原因,怀疑是在神经变性中起作用。随着年龄的增长而上调的感性基因主要参与感测微生物配体的表达,而那些下调的感性基因主要涉及内源性配体的表达[9]。该分析表明神经胶质特异性调节有利于自然神经变性中的神经保护。这与神经退行性疾病中向神经毒性的转变相反。
感觉也可以在神经发育中起作用。小胶质细胞的早期生命感染导致了对后来的免疫刺激高度敏感。当受到感染时,涉及神经炎症的感觉基因上调,而涉及神经可塑性的基因下调。[11]感性的改变神经发育的能力也许能够抵抗疾病。在Rett综合征的啮齿动物模型中删除了高表达的性感基因CX3CL1,从而改善了健康状况并延长了寿命。[12]没有Rett综合征的人中Cx3cr1的下调与类似于精神分裂症的症状有关。[13]这表明,感官不仅在各种发育障碍中起作用,而且还需要严格调节以维持无病状态。
分枝
这种形式的小胶质细胞通常存在于整个大脑和脊髓的特定位置,没有异物或垂死细胞。小胶质细胞的这种“静止”形式由长的分支过程和小的细胞体组成。与小胶质细胞的变形虫形式不同,分支形式的细胞体保留在原处,而其分支不断地移动并测量周围区域。这些分支对生理条件的微小变化非常敏感,需要非常特殊的培养条件才能在体外观察。[8]
与活化的或阿米巴样的小胶质细胞不同,分枝的小胶质细胞不吞噬细胞并且分泌更少的免疫分子(包括MHC I / II类蛋白)。在这种状态下,小胶质细胞能够搜寻和识别免疫威胁,同时维持中枢神经系统的稳态。[14] [15] [16]尽管这被认为是静止状态,但这种形式的小胶质细胞在化学调查环境中仍然非常活跃。分枝的小胶质细胞可以在任何时候因伤害或威胁而转变为活化形式。[8]
反应式(已激活)
尽管历史上经常使用,但“活化的”小胶质细胞应替换为“反应性”的小胶质细胞。[17]确实,显然静止的小胶质细胞并非缺乏活性功能,“激活”一词具有误导性,因为它倾向于指示细胞反应性的“全有或全无”极化。在反应性小胶质细胞中上调的标志物Iba1通常用于可视化这些细胞。[18]
非吞噬
当小胶质细胞从其分支形式转变为完全活跃的吞噬形式时,这种状态实际上是分级反应的一部分。小胶质细胞可以由多种因素激活,包括:促炎性细胞因子,细胞坏死因子,脂多糖和细胞外钾的变化(指示细胞破裂)。活化后,细胞会经历一些关键的形态变化,包括分支的增厚和缩回,MHC I / II类蛋白的摄取,免疫分子的表达,细胞毒性因子的分泌,募集分子的分泌以及促炎性信号分子的分泌(导致在促炎信号级联中)。活化的非吞噬小胶质细胞通常表现为“繁忙”,“杆状”或小类变形虫,这取决于它们分枝到完整的吞噬转化连续体的距离。另外,小胶质细胞也经历快速增殖以增加其数量。从严格的形态学角度来看,沿着连续体的小胶质细胞形式的变化与形态复杂性的变化有关,并且可以使用分形分析的方法进行定量,这种分形分析方法已被证明对不同病理状态下与不同形态相关的微妙的,视觉上无法检测到的变化敏感。[6] [14] [15] [19]
吞噬
活化的吞噬小胶质细胞是小胶质细胞的最大免疫反应形式。尽管已观察到一些差异,但这些细胞通常呈大的类eb形。除了具有活化的非吞噬小胶质细胞的抗原呈递,细胞毒性和炎症介导的信号传导作用外,它们还能够吞噬细胞内的异物并展示出用于T细胞活化的免疫分子。吞噬小胶质细胞到达损伤部位,吞噬令人不快的物质,并分泌促炎因子,以促进更多细胞增殖并进行同样的处理。活化的吞噬小胶质细胞还与星形胶质细胞和神经细胞相互作用,以在不损害健康的脑细胞的情况下,尽快抵抗感染或炎症。[6] [14]
变形虫
这种形状允许小胶质细胞在整个神经组织中自由移动,从而使其能够发挥其作为清道夫细胞的作用。变形虫小胶质细胞能够吞噬细胞碎片,但不能履行与活化小胶质细胞相同的抗原呈递和炎症作用。当有大量细胞外碎片和凋亡细胞要清除时,变形虫小胶质细胞在大脑的发育和重新布线期间尤其普遍。这种小胶质细胞主要存在于被称为“小胶质细胞”的F体的围产期白质区域。[6] [15] [20]
发光细胞
细菌细胞是小胶质细胞吞噬传染性物质或细胞碎片的最终结果。最终,吞噬了一定数量的物质后,吞噬小胶质细胞变得无法吞噬任何其他物质。产生的细胞团称为粒状小球,以其“粒状”外观命名。通过观察染色的组织以揭示细菌的表达,病理学家可以可视化感染后的愈合区域。[21]
血管周围
与上述其他类型的小胶质细胞不同,“血管周围”的小胶质细胞是指细胞的位置,而不是其形式/功能。血管周围小胶质细胞主要发现于基底层壁内。它们具有正常的小胶质细胞功能,但是与正常的小胶质细胞不同,它们会定期被骨髓来源的前体细胞替代,并且无论其环境如何,都表达II类MHC抗原。血管周围小胶质细胞也对巨噬细胞分化抗原产生强烈反应。[6]如Ritter的实验和缺血性视网膜病的观察所表明的,这些小胶质细胞对于血管壁的修复至关重要。血管周围小胶质细胞促进内皮细胞增殖,从而允许形成新的血管并修复受损的血管。在这些过程中,骨髓募集和分化为小胶质细胞的速度大大加快,以完成这些任务。[22]
脉管的
像血管周围小胶质细胞一样,血管旁小胶质细胞主要可以通过其位置来区分。发现脉管小胶质细胞直接接触血管的基底层壁,但在壁内未发现。像血管周细胞一样,它们即使在低水平的炎症细胞因子活性下也表达II类MHC蛋白。与血管周细胞不同,但与常驻小胶质细胞相似,近血管小胶质细胞不表现出快速的更新或定期被髓样前体细胞替代[6]。
功能

通过嘌呤能信号传导激活小胶质细胞
小胶质细胞完成CNS内的各种不同任务,主要与免疫反应和维持体内稳态有关。以下是这些单元执行的一些主要的已知功能。
清除
除了对周围环境的微小变化非常敏感之外,每个小胶质细胞还定期进行物理检查。该动作是在肢体和静止状态下进行的。在小胶质细胞中移动时,如果发现任何异物,受损的细胞,凋亡细胞,神经原纤维缠结,DNA片段或噬菌斑,它将激活并吞噬该物质或细胞。以这种方式,小胶质细胞也可以充当“管家”,清理随机的细胞碎片。[14]在大脑发育过程中,小胶质细胞在调节神经前体细胞数量和清除凋亡神经元方面发挥着重要作用。还有证据表明,小胶质细胞可以通过吞噬和消除突触来改善突触回路。[23]发育后,大多数死亡或凋亡细胞见于大脑皮层和皮层下白质。这也许可以解释为什么在大脑皮层的“小胶质细胞”中发现了大多数类小胶质细胞。[20]
吞噬作用
小胶质细胞的主要作用是吞噬作用,涉及各种物质的吞噬。被吞噬的材料通常由处于非发炎状态的细胞碎片,脂质和凋亡细胞,以及处于发炎状态的入侵病毒,细菌或其他外来物质组成。一旦小胶质细胞“充满”,它将停止吞噬活性,并转变为相对无反应的细菌。
细胞外信号
小胶质细胞在大脑中的作用很大一部分是维持非感染区的稳态,并促进感染或受损组织的炎症。小胶质细胞通过一系列极其复杂的细胞外信号分子来实现这一目标,这些分子可以使它们与其他小胶质细胞,星形胶质细胞,神经,T细胞和骨髓祖细胞进行通讯。如上所述,细胞因子IFN-γ可用于激活小胶质细胞。此外,在被IFN-γ激活后,小胶质细胞还将更多的IFN-γ释放到细胞外空间。这会激活更多的小胶质细胞,并启动细胞因子诱导的激活级联反应,从而迅速激活所有附近的小胶质细胞。小胶质细胞产生的TNF-α使神经组织发生凋亡并增加炎症。 IL-8促进B细胞的生长和分化,从而帮助小胶质细胞抵抗感染。另一种细胞因子IL-1抑制细胞因子IL-10和TGF-β,它们下调抗原引入和促炎信号传导。通过小胶质细胞产生趋化分子(如MDC,IL-8和MIP-3β),将其他树突状细胞和T细胞募集到损伤部位。最后,PGE2和其他类前列腺素可通过抑制小胶质细胞的促炎反应和下调Th1(T辅助细胞)反应来预防慢性炎症。[14]
抗原介绍
如上所述,驻留的未激活的小胶质细胞由于缺乏MHC I / II类蛋白而表现为差的抗原呈递细胞。激活后,它们迅速摄取I / II类MHC蛋白,并迅速成为有效的抗原呈递剂。在某些情况下,小胶质细胞也可以被IFN-γ激活以呈递抗原,但不能像吸收MHC I / II类蛋白质那样有效。在发炎期间,由于特殊的表面标记,T细胞穿过血脑屏障,然后直接与小胶质细胞结合,从而接收抗原。 T细胞一旦获得抗原,便会继续发挥多种作用,包括促炎性募集,免疫记忆的形成,细胞毒性物质的分泌以及对外来细胞质膜的直接攻击。[6] [14]
细胞毒性
除了能够通过吞噬作用通过细胞与细胞的接触破坏感染性生物外,小胶质细胞还可以释放多种细胞毒性物质。培养中的小胶质细胞以“呼吸爆发”的过程分泌大量的过氧化氢和一氧化氮。这两种化学物质都可以直接损伤细胞并导致神经元细胞死亡。小胶质细胞分泌的蛋白酶分解特定蛋白质,导致直接的细胞损伤,而诸如IL-1的细胞因子则促进神经元轴突的脱磷脂磷脂化。最后,小胶质细胞可以通过分泌谷氨酸,天冬氨酸和喹啉酸,通过NMDA受体介导的过程伤害神经元。细胞毒性分泌物旨在破坏受感染的神经元,病毒和细菌,但也可能引起大量附带神经损伤。结果,当小胶质细胞破坏大脑以破坏入侵的感染时,慢性炎症反应可能导致大规模的神经损伤。[6]
突触剥离
1968年,Blinzinger和Kreutzberg首次在脊柱病变中发现这种现象,炎症后小胶质细胞从受损组织附近的神经中去除了分支。这有助于促进受损神经回路的再生长和重新映射。[6]
促进修复
炎症后,小胶质细胞经历了几个步骤来促进神经组织的再生。这些包括突触剥离,抗炎细胞因子的分泌,神经元和星形胶质细胞向受损区域的募集以及金黄色胶质细胞的形成。如果没有小神经胶质细胞,在中枢神经系统的驻留区域中的再生和重新映射将相当慢,而在大脑和眼睛周围的许多血管系统中几乎是不可能的。[6] [22]
发展历程
长期以来,人们认为小胶质细胞在骨髓中与造血干细胞(所有血细胞的祖细胞)相区分。然而,最近的研究表明,小胶质细胞起源于卵黄囊,处于明显受限制的胚胎时期,并分布于脑间质。此外,它们不断更新自我,并在整个生命中持续存在,而没有外周单核细胞前体的补充。[24]
单核细胞还可以在外周系统中分化为髓样树突状细胞和巨噬细胞。像身体其他部位的巨噬细胞一样,小胶质细胞利用吞噬和细胞毒性机制破坏异物。小胶质细胞和巨噬细胞均通过充当抗原呈递细胞来促进免疫反应,并通过分泌细胞因子和其他信号分子促进体内的炎症和体内平衡机制。
小胶质细胞以其下调的形式缺乏I类MHC / II类MHC蛋白,IFN-γ细胞因子,CD45抗原以及许多其他表面受体,这些受体在区分正常巨噬细胞的抗原呈递,吞噬和细胞毒性作用中起作用。小胶质细胞也不同于巨噬细胞,因为它们在空间和时间上受到更严格的调节,以维持精确的免疫反应。[14]
小胶质细胞和其他与髓系祖细胞分化的细胞之间的另一个区别是周转率。巨噬细胞和树突状细胞不断被耗尽,并被分化成所需类型的髓系祖细胞所替代。由于血脑屏障,人体要不断替换小胶质细胞是相当困难的。因此,小胶质细胞不会一直被髓样祖细胞替代,而是保持其静止状态,而当它们被激活时,它们会迅速增殖以保持其数量。然而,骨嵌合体研究表明,在极端感染的情况下,血脑屏障将减弱,小胶质细胞将被血源性骨髓来源的细胞(即骨髓祖细胞和巨噬细胞)取代。一旦感染减少,外围和中央系统之间的连接就会恢复,只有小胶质细胞存在才能恢复和再生。[25]
老化
小胶质细胞在损伤过程中经历有丝分裂活动的爆发;这种增殖之后是凋亡,以减少细胞数量回到基线。[26]小胶质细胞的活化会给细胞的合成代谢和分解代谢机制带来负担,导致活化的小胶质细胞比未活化的细胞更快死亡。[26]为了补偿小胶质细胞随着时间的流失,小胶质细胞经历了有丝分裂,并且骨髓衍生的祖细胞通过脑膜和脉管系统迁移到大脑中。[26]
在正常衰老过程中发生的轻微神经元损伤的积累可将小胶质细胞转化为扩大和活化的细胞。[27]随着年龄的增长,这些慢性,与年龄相关的小胶质细胞激活和IL-1表达的增加可能通过增加易感患者的神经斑块形成而增加了阿尔茨海默氏病的风险。[27] DNA损伤可能与年龄相关的小胶质细胞活化有关。另一个因素可能是随着年龄的增长而积累的高级糖基化终产物。[27]这些蛋白质对蛋白水解过程具有很强的抵抗力,并促进蛋白质交联。[27]
研究发现营养不良(发育不良)的人类小胶质细胞。 “这些细胞的特征在于其胞质结构异常,例如脱枝,萎缩,破碎或异常曲折,经常带有球形或球状肿胀。” [26]营养不良性小胶质细胞的发生率随着年龄的增长而增加。[26]在Prion病,精神分裂症和阿尔茨海默氏病的研究中已经报道了小胶质细胞的变性和死亡,这表明小胶质细胞的退化可能与神经退行性疾病有关。[26]该理论的复杂之处在于,很难区分人脑中的“活化”和“营养不良”小胶质细胞。[26]
在小鼠中,已证明CD22阻断可恢复衰老大脑中的稳态小胶质细胞吞噬作用。[28]

小胶质细胞的图像
临床意义
主要文章:小胶质细胞在疾病中的作用
小胶质细胞是中枢神经系统的主要免疫细胞,类似于外周巨噬细胞。它们通过改变形态并迁移到感染/伤害部位来对病原体和伤害做出反应,在那里他们消灭病原体并清除受损的细胞。作为其反应的一部分,它们分泌细胞因子,趋化因子,前列腺素和活性氧,这有助于指导免疫反应。另外,它们通过产生抗炎细胞因子而有助于炎症反应的解决。小胶质细胞在神经退行性疾病(如阿尔茨海默氏病,帕金森氏病,多发性硬化症以及贲门c病,青光眼以及病毒和细菌感染)中的有害作用也受到了广泛研究。
历史
观察和表征包括小胶质细胞在内的不同神经细胞的能力始于1880年,当时弗朗兹·尼斯尔(Franz Nissl)进行了尼斯尔染色。 Franz Nissl和F.Robertson在组织学实验中首次描述了小胶质细胞。 1880年代的细胞染色技术表明,小胶质细胞与巨噬细胞有关。维克多·贝贝(Victor Babe)在1897年研究狂犬病时首先注意到小胶质细胞的活化和分支的小胶质细胞簇的形成。贝贝(Babe)指出,这些细胞是在各种病毒性脑部感染中发现的,但不知道他所见的小胶质细胞簇是什么。 [29] PíodelRíoHortega是SantiagoRamóny Cajal的学生,他在1920年左右首次将细胞称为“小胶质细胞”。他于1927年继续表征小胶质细胞对脑损伤的反应,并注意到胼胝体和其他围产期中存在的“小胶质细胞”。 1932年成为白质地区。经过多年的研究,里约-霍特加(Rio-Hortega)被普遍认为是“小胶质细胞之父”。[30] [31]长期以来,作者对小胶质细胞的了解几乎没有改善。然后,在1988年,Hickey和Kimura表明血管周微胶质细胞是骨髓来源的,并表达高水平的MHC II类蛋白质用于抗原引入。这证实了Pio Del Rio-Hortega的假设,即小胶质细胞通过执行吞噬作用和抗原引入来发挥与巨噬细胞相似的功能。
另见
Astrocyte
Macrophage
Neuroimmune system
List of human cell 类型 derived from the germ layers
参考
Ginhoux F, Lim S, Hoeffel G, Low D, Huber T (2013). "Origin and differentiation of microglia". Frontiers in Cellular Neuroscience. 7: 45. doi:10.3389/fncel.2013.00045. PMC 3627983. PMID 23616747.
Lawson LJ, Perry VH, Gordon S (1992). "Turnover of resident microglia in the normal adult mouse brain". Neuroscience. 48 (2): 405–15. doi:10.1016/0306-4522(92)90500-2. PMID 1603325.
Filiano AJ, Gadani SP, Kipnis J (August 2015). "Interactions of innate and adaptive immunity in brain development and function". Brain Research. 1617: 18–27. doi:10.1016/j.brainres.2014.07.050. PMC 4320678. PMID 25110235.
Kreutzberg GW (March 1995). "Microglia, the first line of defence in brain pathologies". Arzneimittel-Forschung. 45 (3A): 357–60. PMID 7763326.
Bushong EA, Martone ME, Jones YZ, Ellisman MH (January 2002). "Protoplasmic astrocytes in CA1 stratum radiatum occupy separate anatomical domains". The Journal of Neuroscience. 22 (1): 183–92. doi:10.1523/JNEUROSCI.22-01-00183.2002. PMID 11756501.
Gehrmann J, Matsumoto Y, Kreutzberg GW (March 1995). "Microglia: intrinsic immuneffector cell of the brain". Brain Research. Brain Research Reviews. 20 (3): 269–87. doi:10.1016/0165-0173(94)00015-H. PMID 7550361.
Dissing-Olesen L, Ladeby R, Nielsen HH, Toft-Hansen H, Dalmau I, Finsen B (October 2007). "Axonal lesion-induced microglial proliferation and microglial cluster formation in the mouse". Neuroscience. 149 (1): 112–22. doi:10.1016/j.neuroscience.2007.06.037. PMID 17870248.
Verkhratsky A, Butt A (2013). Glial physiology and pathophysiology. Chicester: John Wiley & Sons. ISBN 978-1118402054.[page needed]
Hickman SE, Kingery ND, Ohsumi TK, Borowsky ML, Wang LC, Means TK, El Khoury J (2013). "The microglial sensome revealed by direct RNA sequencing". Nature Neuroscience. 16 (12): 1896–905. doi:10.1038/nn.3554. PMC 3840123. PMID 24162652.
Block, M.L., Zecca, L. & Hong, J.S. Microglia-mediated neurotoxicity: uncovering the molecular mechanisms. Nat. Rev. Neurosci. 8, 57–69 (2007).
Ji P, Schachtschneider KM, Schook LB, Walker FR, Johnson RW (2016). "Peripheral viral infection induced microglial sensome genes and enhanced microglial cell activity in the 海马 of neonatal piglets". Brain, Behavior, and Immunity. 54: 243–251. doi:10.1016/j.bbi.2016.02.010. PMC 4828316. PMID 26872419.
Horiuchi M, Smith L, Maezawa I, Jin LW (2017). "CX3CR1 ablation ameliorates motor and respiratory dysfunctions and improves survival of a Rett syndrome mouse model". Brain, Behavior, and Immunity. 60: 106–116. doi:10.1016/j.bbi.2016.02.014. PMC 5531048. PMID 26883520.
Bergon A, Belzeaux R, Comte M, Pelletier F, Hervé M, Gardiner EJ, et al. (2015). "CX3CR1 is dysregulated in blood and brain from schizophrenia 病人" (PDF). Schizophrenia Research. 168 (1–2): 434–43. doi:10.1016/j.schres.2015.08.010. PMID 26285829.
Aloisi F (November 2001). "Immune function of microglia". Glia. 36 (2): 165–79. doi:10.1002/glia.1106. PMID 11596125.
Christensen RN, Ha BK, Sun F, Bresnahan JC, Beattie MS (July 2006). "Kainate induces rapid redistribution of the actin cytoskeleton in ameboid microglia". Journal of Neuroscience Research. 84 (1): 170–81. doi:10.1002/jnr.20865. PMID 16625662.
Davis EJ, Foster TD, Thomas WE (1994). "Cellular forms and functions of brain microglia". Brain Research Bulletin. 34 (1): 73–8. doi:10.1016/0361-9230(94)90189-9. PMID 8193937.
Eggen BJ, Raj D, Hanisch UK, Boddeke HW (September 2013). "Microglial phenotype and adaptation". Journal of Neuroimmune Pharmacology. 8 (4): 807–23. doi:10.1007/s11481-013-9490-4. PMID 23881706.
Lan X, Han X, Li Q, Yang QW, Wang J (July 2017). "Modulators of microglial activation and polarization after intracerebral haemorrhage". Nat Rev Neurol. 13 (7): 420–433. doi:10.1038/nrneurol.2017.69. PMC 5575938. PMID 28524175.
Jelinek HF, Karperien A, Bossomaier T, Buchan A (1975). "Differentiating grades of microglia activation with fractal analysis" (PDF). Complexity International. 12 (18): 1713–7. Archived from the original (PDF) on 2008-12-17.
Ferrer I, Bernet E, Soriano E, del Rio T, Fonseca M (1990). "Naturally occurring cell death in the cerebral cortex of the rat and removal of dead cells by transitory phagocytes". Neuroscience. 39 (2): 451–8. doi:10.1016/0306-4522(90)90281-8. PMID 2087266.
Rissi DR, Oliveira FN, Rech RR, Pierezan F, Lemos RA, Barros CS (2006). "Epidemiologia, sinais clínicos e distribuio das lesões encefálicas em bovinos afetados por meningoencefalite por herpesvírus bovino-5" [Epidemiology, clinical signs and distribution of the encephalic lesions in cattle affected by meningoencephalitis caused by bovine herpesvirus-5]. Pesquisa Veterinária Brasileira (in Portuguese). 26 (2): 123–32. doi:10.1590/S0100-736X2006000200010.
Ritter MR, Banin E, Moreno SK, Aguilar E, Dorrell MI, Friedlander M (December 2006). "Myeloid progenitors differentiate into microglia and promote vascular 修复 in a model of ischemic retinopathy". The Journal of Clinical Investigation. 116 (12): 3266–76. doi:10.1172/JCI29683. PMC 1636693. PMID 17111048.
Chung WS, Welsh CA, Barres BA, Stevens B (November 2015). "Do glia drive synaptic and cognitive impairment in disease?". Nature Neuroscience. 18 (11): 1539–1545. doi:10.1038/nn.4142. PMC 4739631. PMID 26505565.
Ginhoux F, Prinz M (July 2015). "Origin of microglia: current concepts and past controversies". Cold Spring Harbor Perspectives in Biology. 7 (8): a020537. doi:10.1101/cshperspect.a020537. PMC 4526747. PMID 26134003.
Gehrmann J (1996). "Microglia: a sensor to threats in the nervous system?". Research in Virology. 147 (2–3): 79–88. doi:10.1016/0923-2516(96)80220-2. PMID 8901425.
Streit WJ (September 2006). "Microglial senescence: does the brain's immune system have an expiration date?". Trends in Neurosciences. 29 (9): 506–10. doi:10.1016/j.tins.2006.07.001. PMID 16859761.
Mrak RE, Griffin WS (March 2005). "Glia and their cytokines in progression of neurodegeneration". Neurobiology of Aging. 26 (3): 349–54. doi:10.1016/j.neurobiolaging.2004.05.010. PMID 15639313.
Pluvinage JV, Wyss-Coray T, et al. (April 11, 2019). "CD22 blockade restores homeostatic microglial phagocytosis in aging brains". Nature. 568 (7751): 187–192. doi:10.1038/s41586-019-1088-4.
Babeş, VM (1892). "Certains caractères des lesions histologiques de la rage" [Certain characteristics of the histological lesions of rabies]. Annales de l'Institut Pasteur (in French). 6: 209–23.
del Río Hortega P, Penfield W (1892). "Cerebral Cicatrix: the Reaction of Neuroglia and Microglia to Brain Wounds". Bulletin of the Johns Hopkins Hospital. 41: 278–303.
del Rio-Hortega F (1937). "Microglia". Cytology and Cellular Pathology of the Nervous System: 481–534. |

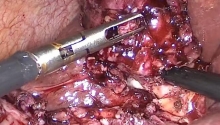
 急性胆囊炎——紧急救治策略
急性胆囊炎——紧急救治策略
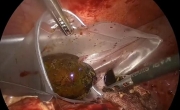
 采用腹腔镜经筋膜腹外缝合术修复因 Morgagn
采用腹腔镜经筋膜腹外缝合术修复因 Morgagn
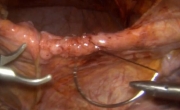
 腹腔镜全胃切除术治疗胃癌中采用线性吻合器
腹腔镜全胃切除术治疗胃癌中采用线性吻合器
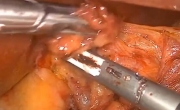
 腹腔镜下Whipple手术治疗胰头肿瘤
腹腔镜下Whipple手术治疗胰头肿瘤

 发表于 2019-12-21 00:00:24
发表于 2019-12-21 00:00:24
 提升卡
提升卡 置顶卡
置顶卡 沉默卡
沉默卡 喧嚣卡
喧嚣卡 变色卡
变色卡 千斤顶
千斤顶 显身卡
显身卡