马上注册,结交更多好友,享用更多功能,让你轻松玩转社区。
您需要 登录 才可以下载或查看,没有账号?注册
×
星形胶质细胞(来自希腊语astron的Astro =星形,来自希腊语“ kytos”的cyte表示腔),也统称为星形胶质细胞,是大脑和脊髓中特征性的星形胶质细胞。大脑中星形胶质细胞的比例尚不明确。根据所使用的计数技术,研究发现,星形胶质细胞的比例因区域而异,占所有神经胶质细胞的20%至40%。[1]它们执行许多功能,包括形成血脑屏障的内皮细胞的生化支持,向神经组织提供营养,维持细胞外离子平衡以及在创伤后脑和脊髓的修复和疤痕形成过程中发挥作用。抽动受伤。
自1990年代中期以来的研究表明,星形胶质细胞响应刺激而长距离传播细胞间Ca2 +波,并且类似于神经元,以Ca2 +依赖的方式释放递质(称为神经胶质递质)。[2]数据表明,星形胶质细胞还通过钙离子依赖性谷氨酸的释放向神经元发出信号。[3]这些发现使星形胶质细胞成为神经科学领域的重要研究领域。

来自大鼠脑的星形胶质细胞在组织培养物中生长,并用针对GFAP(红色)和波形蛋白(绿色)的抗体染色。 两种蛋白质均大量存在于该细胞的中间丝中,因此细胞呈黄色。 蓝色物质显示用DAPI染色剂可视化的DNA,并显示星形胶质细胞和其他细胞的细胞核。 图片由EnCor Biotechnology Inc.提供
内容
1 结构
1.1 发展
2 功能
2.1 三方突触
3 临床意义
3.1 星状细胞瘤
3.2 神经发育障碍
3.3 慢性疼痛
3.3.1 持续疼痛的两种状态
3.4 其他病理
4 研究
5 分类
5.1 谱系和抗原表型
5.2 解剖学分类
5.3 转运蛋白/受体的分类
6 参考
结构
 in the context of neurons (red) in a mouse cortex cell culture.jpg")
小鼠皮质细胞培养物中神经元(红色)中的星形胶质细胞(绿色)

23周龄胎儿大脑培养人星形胶质细胞
 among neurons (green) in the living cerebral cortex.png")
活的大脑皮层神经元(绿色)中的星形胶质细胞(红黄色)
星形胶质细胞是中枢神经系统中神经胶质细胞的一种亚型。它们也被称为星形胶质细胞。呈星形,它们的许多过程都包裹着神经元产生的突触。传统上使用组织学分析鉴定星形胶质细胞。这些细胞中的许多表达中间丝状胶质纤维酸性蛋白(GFAP)。[4]中枢神经系统中存在几种形式的星形胶质细胞,包括纤维状(白质),原生质体(灰质)和桡骨。纤维状神经胶质细胞通常位于白质内,细胞器相对较少,并表现出长而无分支的细胞过程。这种类型通常具有“血管脚”,当它们靠近毛细管壁时,它们会将细胞物理连接到毛细管壁的外部。原生质胶质细胞最普遍,见于灰质组织中,具有大量的细胞器,并表现出短而高度分支的三级过程。桡骨胶质细胞位于垂直于心室轴的平面中。他们的一个过程毗邻软脑膜,而另一个过程则深深地埋在灰质中。桡骨胶质细胞主要在发育过程中出现,在神经元迁移中起作用。视网膜的Müller细胞和小脑皮层的Bergmann胶质细胞是一个例外,在成年期仍然存在。当接近软脑膜时,所有三种形式的星形胶质细胞都发出形成皮神经胶质膜的过程。
发展历程

星形胶质细胞以红色表示。细胞核以蓝色表示。从新生小鼠的大脑中获得星形胶质细胞
星形胶质细胞是中枢神经系统中的大胶质细胞。星形胶质细胞来自发育中枢神经系统神经上皮细胞的祖细胞异质群体。在指定多种神经元亚型谱系和大胶质细胞谱系的众所周知的遗传机制之间存在着显著的相似性。[5]就像神经元细胞规范一样,诸如Sonic刺猬(SHH),成纤维细胞生长因子(FGFs),WNT和骨形态发生蛋白(BMP)的规范性信号传导因子通过沿背-腹,前,前的形态发生因子梯度为发展大胶质细胞提供位置信息后轴和内侧轴。沿着神经轴形成的图案导致神经上皮细分为发育脊髓中不同神经元类型的祖域(p0,p1,p2,p3和pMN)。根据一些研究,现在认为该模型也适用于大胶质细胞规范。 Hochstim及其同事进行的研究表明,p1,p2和p3结构域产生了三个不同的星形胶质细胞群体。[6]星形胶质细胞的这些亚型可以根据它们在不同转录因子(PAX6,NKX6.1)和细胞表面标志物(reelin和SLIT1)的表达来确定。已鉴定出的三个星形细胞亚型群体是:1)背侧位于VA1星形胶质细胞,源自p1结构域,表达PAX6和reelin 2)背侧位于VA3星形胶质细胞,源自p3,表达NKX6.1和SLIT1和3)和中间位于p2域的白色物质VA2星形胶质细胞,表达PAX6,NKX6.1,reelin和SLIT1。[7]在发育中枢神经系统中出现星形胶质细胞规格后,据信星形胶质细胞前体在发生终末分化过程之前迁移至神经系统内的最终位置。
功能
![Metabolic interactions between astrocytes and neurons.[8].png](data/attachment/forum/201911/12/121146nwgccgc4wkr4gmdc.png "Metabolic interactions between astrocytes and neurons.[8].png")
星形胶质细胞和神经元之间的代谢相互作用。[8]
星形胶质细胞帮助形成大脑的物理结构,并被认为发挥许多积极作用,包括神经递质的分泌或吸收以及血脑屏障的维持。[9]提出了三重突触的概念,指的是突触前元件,突触后元件和神经胶质元件之间在突触中发生的紧密关系。[10]
结构:它们参与大脑的物理结构。星状细胞之所以得名,是因为它们是“星形”的。它们是大脑中与神经元突触密切相关的最丰富的神经胶质细胞。它们调节脑内电脉冲的传输。
糖原燃料储备缓冲液:星形胶质细胞含有糖原,能够糖异生。额叶皮层和海马中神经元旁边的星形胶质细胞储存并释放葡萄糖。因此,在高葡萄糖消耗率和葡萄糖短缺期间,星形胶质细胞可以为神经元提供葡萄糖。最近对大鼠的研究表明,这种活动与体育锻炼之间可能存在联系。[11]
代谢支持:它们为神经元提供营养,例如乳酸。
葡萄糖感测:通常与神经元有关,大脑中间质葡萄糖水平的检测也由星形胶质细胞控制。星形胶质细胞在体外被低糖激活,在体内这种激活会增加胃排空以增加消化。[12]
血脑屏障:围绕星形胶质细胞末端的内皮细胞被认为有助于维持血脑屏障,但最近的研究表明它们并未发挥实质性作用。相反,在维持屏障中最重要的作用是大脑内皮细胞的紧密连接和基底层。[13]然而,最近发现星形胶质细胞的活性与大脑中的血流有关,而这正是功能磁共振成像所实际测量的。[14] [15]
递质的摄取和释放:星形胶质细胞表达质膜转运蛋白,例如谷氨酸转运蛋白,用于多种神经递质,包括谷氨酸,ATP和GABA。最近,显示星形胶质细胞以囊泡,Ca2 +依赖性的方式释放谷氨酸或ATP。[16] (这已被海马星形胶质细胞争议。)[17]
细胞外空间离子浓度的调节:星形胶质细胞以高密度表达钾通道。当神经元活跃时,它们会释放钾离子,从而增加局部细胞外浓度。由于星形胶质细胞对钾具有很高的渗透性,因此它们可以迅速清除细胞外空间中的过量积累。[18]如果该功能受到干扰,钾的细胞外浓度将升高,从而通过高盛方程导致神经元去极化。众所周知,细胞外钾的异常积累会导致癫痫神经元活动。[19]
突触传递的调节:在丘脑的视上核中,星形胶质细胞形态的快速变化已显示出会影响神经元之间的异突触传递。[20]在海马中,星形胶质细胞通过释放ATP抑制突触传递,而ATP被外切核苷酸酶水解产生腺苷。腺苷作用于神经元腺苷受体以抑制突触传递,从而增加了LTP的动态范围。[21]
血管调节:星形胶质细胞可能是神经元调节血流的中介。[22]
促进少突胶质细胞的脊髓磷脂化活性的增强:神经元的电活动使它们释放ATP,这是形成髓磷脂的重要刺激因素。但是,ATP并不直接作用于少突胶质细胞。相反,它使星形胶质细胞分泌细胞因子白血病抑制因子(LIF),一种调节蛋白,可促进少突胶质细胞的髓磷脂形成活性。这表明星形胶质细胞在大脑中具有执行协调作用。[23]
神经系统修复:损伤中枢神经系统中的神经细胞后,星形胶质细胞会填满整个空间,形成神经胶质瘢痕,并可能有助于神经修复。星形胶质细胞在损伤后中枢神经系统再生中的作用尚不清楚。传统上将神经胶质瘢痕描述为再生的不可渗透的屏障,因此暗示了轴突再生的负面作用。然而,最近,通过基因消融研究发现,星形胶质细胞实际上是再生发生所必需的。[24]更重要的是,作者发现星形胶质瘢痕实际上对于刺激的轴突(通过神经营养补充被诱使轴突生长)延伸穿过受伤的脊髓至关重要。[24]已被推入反应性表型的星形胶质细胞(称为星形胶质细胞增生症,由上调GFAP表达定义,这一定义仍在争论中)实际上可能对神经元有毒,释放出可以杀死神经元的信号。[25]然而,仍有许多工作来阐明它们在神经系统损伤中的作用。
长期增强作用:科学家们争论星形胶质细胞是否整合海马中的学习和记忆能力。 最近已经显示出,将人神经胶质祖细胞移植到新生小鼠的大脑中会导致该细胞分化为星形胶质细胞。 分化后,这些细胞增加了LTP并改善了小鼠的记忆力。[26]
昼夜节律时钟:仅星形胶质细胞就足以驱动SCN中的分子振荡和小鼠的昼夜节律行为,因此可以自主启动和维持复杂的哺乳动物行为。[27]
神经系统的转换:基于以下证据,[28]最近有人推测,大神经胶质细胞(尤其是星形胶质细胞)既是有损神经递质的电容器,又是神经系统的逻辑转换。 即,大胶质细胞根据其膜状态和刺激水平阻止或使刺激沿着神经系统传播。

图6如Nossenson等[28] [29]所暗示的,胶质细胞在生物神经检测方案中的推测转换作用。
Ca2 +离子流入星形胶质细胞是必不可少的变化,最终会产生钙波。由于这种流入是直接由流向大脑的血液增加引起的,因此钙波被认为是一种血液动力学反应功能。细胞内钙浓度的增加可通过该功能合胞体向外传播。钙波传播的机制包括钙离子和IP3通过间隙连接和细胞外ATP信号传导的扩散。[39]钙升高是星形胶质细胞激活的主要已知轴,对于某些类型的星形细胞谷氨酸释放来说,钙的升高是必要和充分的。[40]考虑到钙信号在星形胶质细胞中的重要性,已经开发了用于时空钙信号发展的严格调控机制。通过数学分析表明,Ca2 +离子的局部流入会导致Ca2 +离子的胞质浓度局部升高。[41]此外,胞质中Ca2 +的积累与每个细胞内钙通量无关,并且取决于跨膜的Ca2 +交换,胞质钙的扩散,细胞的几何形状,细胞外钙的摄动和初始浓度。[41]
三方突触
主条目:三方突触
在脊髓背角内,活化的星形胶质细胞具有对几乎所有神经递质作出反应的能力[42],并在活化后释放多种神经活性分子,例如谷氨酸,ATP,一氧化氮(NO)和前列腺素(PG)。 ),进而影响神经元的兴奋性。星形胶质细胞与突触前和突触后末端之间的紧密联系,以及它们整合突触活性和释放神经调节剂的能力,被称为三方突触。[10]由于这种三部分的结合,星形胶质细胞发生突触调节。
临床意义
星状细胞瘤
星形细胞瘤是从星形胶质细胞发展而来的原发性颅内肿瘤。神经胶质祖细胞或神经干细胞也可能引起星形细胞瘤。这些肿瘤可能发生在大脑和/或脊髓的许多部位。星形细胞瘤分为两类:低度(I和II)和高度(III和IV)。低度肿瘤在儿童中更为常见,而高度肿瘤在成人中更为常见。恶性星形细胞瘤在男性中更为普遍,导致生存率下降。[43]
上皮星形细胞瘤是I级肿瘤。它们被认为是良性和缓慢生长的肿瘤。上叶星形细胞瘤通常具有充满液体的结节部分和结节,结节是固体部分。大部分位于小脑。因此,大多数症状与平衡或协调困难有关。[43]它们在儿童和青少年中也更常见。[44]
纤维状星形细胞瘤是II级肿瘤。它们生长相对较慢,因此通常被认为是良性的,但它们会渗入周围的健康组织并可能变得恶性。纤维状星形细胞瘤通常发生在年轻人中,这些年轻人经常发作。[44]
间变性星形细胞瘤是Ⅲ级恶性肿瘤。它们比低等级的肿瘤生长更快。间变性星形细胞瘤的复发率比低度恶性肿瘤高,因为它们扩散到周围组织的趋势使其很难通过手术彻底清除。[43]
多形胶质母细胞瘤是IV级癌症,可能起源于星形胶质细胞或现有的星形细胞瘤。所有脑肿瘤中约有50%是胶质母细胞瘤。胶质母细胞瘤可包含多种胶质细胞类型,包括星形胶质细胞和少突胶质细胞。胶质母细胞瘤生长迅速并扩散到附近组织,因此通常被认为是最具侵袭性的神经胶质瘤类型。治疗可能很复杂,因为一种肿瘤细胞类型可能会因特定治疗而死亡,而另一种细胞类型可能会继续繁殖。[43]
神经发育障碍
近年来,星形胶质细胞已成为各种神经发育疾病过程的重要参与者。这种观点认为,星形胶质细胞功能障碍可能导致神经回路异常,这是某些精神疾病的基础,例如自闭症谱系障碍(ASD)和精神分裂症。[45] [46]
慢性疼痛
在正常情况下,疼痛传导从某种有害信号开始,然后是伤害性(疼痛感)传入神经元所携带的动作电位,在脊髓背角引起兴奋性突触后电位(EPSP)。然后,该消息将传递到大脑皮层,在此处作者将这些EPSP转换为“疼痛”。自从发现星形胶质细胞-神经元信号传导以来,作者对疼痛传导的理解就变得极为复杂。疼痛处理不再被看作是从身体到大脑的信号的重复中继,而是被视为一个复杂的系统,可以通过许多不同的因素来上下调节。最近研究的最前沿因素之一是位于脊髓背角的增强疼痛的突触以及星形胶质细胞在封装这些突触中的作用。 Garrison和同事[47]在发现脊髓背角的星形胶质细胞肥大与周围神经损伤后对疼痛的超敏反应(通常被认为是损伤后神经胶质活化的指标)之间的相关性时,首先提出了关联。星形胶质细胞检测神经元活性并释放化学递质,进而控制突触活性。[42] [48] [49]过去,痛觉过敏被认为是由P物质和兴奋性氨基酸(EAA)(如谷氨酸)从脊髓背角突触前传入神经末梢的释放所调节的。随后激活了离子型谷氨酸受体的AMPA(α-氨基-3-羟基-5-甲基-4-异恶唑丙酸),NMDA(N-甲基-D-天冬氨酸)和海藻酸酯亚型。这些受体的激活增强了脊髓上的疼痛信号。这个想法虽然是正确的,但却是对疼痛转导的过度简化。合成了一系列其他神经递质和神经调节剂,例如降钙素基因相关肽(CGRP),三磷酸腺苷(ATP),脑源性神经营养因子(BDNF),生长抑素,血管活性肠肽(VIP),甘丙肽和血管加压素并释放出来以响应有害刺激。除了这些调节因子之外,疼痛传递神经元与背角中其他神经元之间的其他几种相互作用也增加了对疼痛途径的影响。
持续疼痛的两种状态
持续的周围组织损伤后,受伤组织以及脊髓背角会释放多种因子。这些因素增加了背角疼痛投射神经元对随之而来的刺激的反应性,称为“脊髓敏化”,从而放大了对大脑的疼痛冲动。谷氨酸,P物质和降钙素基因相关肽(CGRP)的释放介导NMDAR激活(最初是沉默的,因为它被Mg2 +堵住了),从而有助于突触后疼痛传递神经元(PTN)的去极化。此外,IP3信号传导和MAPKs(促分裂原激活的蛋白激酶)(如ERK和JNK)的激活导致改变谷氨酸转运蛋白功能的炎症因子的合成增加。 ERK还可以激活神经元中的AMPAR和NMDAR。 ATP和P物质与它们各自的受体(P2X3)和神经激肽1受体(NK1R)的缔合,以及代谢型谷氨酸受体的活化和BDNF的释放,进一步使伤害感受变得敏锐。谷氨酸持续存在于突触中,最终导致谷氨酸进入星形胶质细胞的重要转运蛋白GLT1和GLAST的失调。持续的兴奋还可以诱导ERK和JNK活化,从而导致多种炎症因子的释放。
随着伤害性疼痛的持续,脊柱敏化会在背角神经元中产生转录变化,从而导致功能改变延长。内部贮存中Ca2 +的操作结果来自持续的突触活动,并导致释放谷氨酸,ATP,肿瘤坏死因子-α(TNF-α),白介素1β(IL-1β),IL-6,一氧化氮(NO),和前列腺素E2(PGE2)。活化的星形胶质细胞也是基质金属蛋白酶2(MMP2)的来源,该酶诱导前IL-1β裂解并维持星形胶质细胞活化。在该慢性信号传导途径中,IL-1β信号传导激活了p38,并且存在趋化因子触发其受体变得活跃。响应神经损伤,热休克蛋白(HSP)被释放并可以与其各自的TLR结合,从而导致进一步的激活。
其他病理
另见:神经免疫系统
涉及星形胶质细胞的其他临床上重要的病理学包括星形胶质细胞增生和星形细胞病变。这些例子包括多发性硬化症,抗AQP4 +视神经脊髓炎,拉斯穆森氏脑炎,亚历山大氏病和肌萎缩性侧索硬化症。[50]研究表明,星形胶质细胞可能隐含在神经退行性疾病中,例如阿尔茨海默氏病,[51] [52]帕金森氏病,[53]亨廷顿氏病和阿米巴侧索硬化症。[54]
研究
罗彻斯特大学和科罗拉多大学医学院的一组科学家于2010年11月进行了一项研究,并于2011年3月发表了这项研究。他们进行了一项实验,试图通过替换神经胶质细胞来修复成年大鼠的中枢神经系统的损伤。当将神经胶质细胞注入成年大鼠脊髓的损伤中时,星形胶质细胞是通过将人类神经胶质前体细胞暴露于骨形态发生蛋白(骨形态发生蛋白很重要,因为它被认为可以在整个人体中形成组织结构)而产生。因此,结合骨蛋白和人类神经胶质细胞,它们促进了明显的足部位置恢复,轴突生长以及脊髓板层神经元存活的明显增加。另一方面,人神经胶质前体细胞和这些细胞通过与睫状神经营养因子接触而产生的星形胶质细胞未能促进神经元存活并在损伤部位促进轴突生长。[55]
在上海进行的一项研究有两种类型的海马神经元培养物:一种培养物中,神经元是从一层星形胶质细胞生长而来的,另一种培养物则不与任何星形胶质细胞接触,而是用神经胶质条件培养基(GCM)喂养在大多数情况下,它会抑制培养的星形胶质细胞在大鼠大脑中的快速生长。在他们的结果中,他们能够看到,星形胶质细胞在混合培养物(是从星形胶质细胞层中生长的培养物)的长期增强中具有直接作用,而在GCM培养物中则没有。[56]
研究表明,星形胶质细胞在神经干细胞的调节中起重要作用。哈佛大学Schepens眼研究所的研究表明,人脑中神经干细胞大量存在,神经干细胞通过来自星形胶质细胞的化学信号(ephrin-A2和ephrin-A3)保持休眠状态。星形胶质细胞能够通过抑制ephrin-A2和ephrin-A3的释放来激活干细胞转化为工作神经元。[57]
威斯康星大学的一组研究人员在2011年《自然生物技术》杂志上发表的一项研究[58]中报告说,它已经能够指导胚胎干细胞和诱导的人类干细胞成为星形胶质细胞。
2012年对大麻对短期记忆的影响的研究[59]发现,四氢大麻酚激活星形胶质细胞的CB1受体,从而导致AMPA受体从相关神经元的膜上被去除。
分类
有几种不同的方法来分类星形胶质细胞。
谱系和抗原表型
这些已经由Raff等人的经典著作确立。在1980年代初期使用Rat视神经。
1型:抗原性Ran2 +,GFAP +,FGFR3 +,A2B5,因此类似于出生后第7天大鼠视神经的“ 1型星形胶质细胞”。这些可能来自三能神经胶质细胞限制性前体细胞(GRP),而不是双能O2A / OPC(少突胶质细胞,2型星形胶质细胞前体,也称为少突胶质细胞祖细胞)细胞。
类型2:抗原性A2B5 +,GFAP +,FGFR3,Ran2。这些细胞可以从三能GRP(可能通过O2A阶段)或双能O2A细胞(有人{{[60]}}认为可能反过来)体外发育。这些祖细胞被移植到病变部位(但可能不是正常发育,至少不是在大鼠视神经中)时,它们已经衍生自GRP)或体内。 2型星形胶质细胞是产后视神经培养物中主要的星形细胞成分,是由胎牛血清存在下生长的O2A细胞产生的,但认为在体内不存在。[61]
解剖分类
原生质:在灰质中发现,有许多分支过程,其末端包裹着突触。多能性脑室下区祖细胞产生一些原生质体星形胶质细胞。[62] [63]
Gmri阳性星形胶质细胞。这些是原生质体星形胶质细胞的子集,其中包含许多胞质内含物或颗粒,它们被Gmri的铬-铝苏木精染色呈阳性。现已知道,这些颗粒是由溶酶体内吞噬的变性线粒体的残余物形成的。[64]某些类型的氧化应激似乎是这些特殊星形胶质细胞内线粒体损伤的原因。在丘丘脑弓形核和海马中,Gmri阳性星形胶质细胞比其他脑区域丰富得多。它们可能在调节次丘脑对葡萄糖的反应中起作用。[65] [66]
纤维状的:存在于白质中,具有长而细的未分支过程,其末端脚包围兰维尔的结节。 [67] [68] [69] [70] [71]由桡骨胶质细胞产生一些纤维状星形胶质细胞。
转运蛋白/受体分类
GluT类型:这些表达谷氨酸转运蛋白(EAAT1 / SLC1A3和EAAT2 / SLC1A2),并通过转运蛋白电流响应谷氨酸的突触释放。 EAAT2的功能和可用性受人类星形胶质细胞中的细胞内受体TAAR1调节。[72]
GluR类型:这些表达谷氨酸受体(主要是mGluR和AMPA类型),并通过通道介导的电流和IP3依赖性Ca2 +瞬变来响应谷氨酸的突触释放。
另见
伯格曼神经胶质病
生殖细胞
桡骨胶质细胞
神经胶质
垂体细胞
聚树突状细胞
源自细菌层的人类细胞类型列表
参考
Verkhratsky A, Butt AM (2013). "Numbers: how many glial cells are in the brain?". Glial Physiology and Pathophysiology. John Wiley and Sons. pp. 93–96. ISBN 978-0-470-97853-5.
"Role of Astrocytes in the Central Nervous System". Retrieved 27 July 2018.
Fiacco TA, Agulhon C, McCarthy KD (October 2008). "Sorting out astrocyte physiology from pharmacology". Annual Review of Pharmacology and Toxicology. 49 (1): 151–74. doi:10.1146/annurev.pharmtox.011008.145602. PMID 18834310.
Venkatesh K, Srikanth L, Venγ B, Chandrasekhar C, Sanjeevkumar A, Mouleshwara Prasad BC, Sarma PV (2013). "In vitro differentiation of cultured human CD34+ cells into astrocytes". Neurology India. 61 (4): 383–8. doi:10.4103/0028-3886.117615. PMID 24005729.
Rowitch DH, Kriegstein AR (November 2010). "Developmental genetics of vertebrate glial-cell specification". Nature. 468 (7321): 214–22. Bibcode:2010Natur.468..214R. doi:10.1038/nature09611. PMID 21068830.
Muroyama Y, Fujiwara Y, Orkin SH, Rowitch DH (November 2005). "Specification of astrocytes by bHLH protein SCL in a restricted region of the neural tube". Nature. 438 (7066): 360–3. Bibcode:2005Natur.438..360M. doi:10.1038/nature04139. PMID 16292311.
Hochstim C, Deneen B, Lukaszewicz A, Zhou Q, Anderson DJ (May 2008). "Identification of positionally distinct astrocyte sub类型 whose identities are specified by a homeodomain code". Cell. 133 (3): 510–22. doi:10.1016/j.cell.2008.02.046. PMC 2394859. PMID 18455991.
Cakir T, Alsan S, Saybaşili H, Akin A, Ulgen KO (December 2007). "Reconstruction and flux analysis of coupling between metabolic pathways of astrocytes and neurons: application to cerebral hypoxia". Theoretical Biology & Medical Modelling. 4 (1): 48. doi:10.1186/1742-4682-4-48. PMC 2246127. PMID 18070347.
Kolb, Brian and Whishaw, Ian Q. (2008) Fundamentals of Human Neuropsychology. Worth Publishers. 6th ed. ISBN 0716795868
Araque A, Parpura V, Sanzgiri RP, Haydon PG (May 1999). "Tripartite synapses: glia, the unacknowledged partner". Trends in Neurosciences. 22 (5): 208–15. doi:10.1016/S0166-2236(98)01349-6. PMID 10322493.
Reynolds, Gretchen (22 February 2012). "How Exercise Fuels the Brain". New York Times.
McDougal DH, Viard E, Hermann GE, Rogers RC (April 2013). "Astrocytes in the hindbrain detect glucoprivation and regulate gastric motility". Autonomic Neuroscience. 175 (1–2): 61–9. doi:10.1016/j.autneu.2012.12.006. PMC 3951246. PMID 23313342.
Kimelberg HK, Jalonen T, Walz W (1993). "Regulation of the brain microenvironment:transmitters and ions.". In Murphy S (ed.). Astrocytes: pharmacology and function. San Diego, CA: Academic Press. pp. 193–222. ISBN 978-0125113700.
Swaminathan N (1 October 2008). "Brain-scan mystery solved". Scientific American Mind: 7.
Figley CR, Stroman PW (February 2011). "The role(s) of astrocytes and astrocyte activity in neurometabolism, neurovascular coupling, and the production of functional neuroimaging signals". The European Journal of Neuroscience. 33 (4): 577–88. doi:10.1111/j.1460-9568.2010.07584.x. PMID 21314846.
Santello M, Volterra A (January 2009). "Synaptic modulation by astrocytes via Ca2+-dependent glutamate release". Neuroscience. Mar. 158 (1): 253–9. doi:10.1016/j.neuroscience.2008.03.039. PMID 18455880.
Agulhon C, Fiacco TA, McCarthy KD (March 2010). "Hippocampal short- and long-term plasticity are not modulated by astrocyte Ca2+ signaling". Science. 327 (5970): 1250–4. Bibcode:2010Sci...327.1250A. doi:10.1126/science.1184821. PMID 20203048.
Walz W (April 2000). "Role of astrocytes in the clearance of excess extracellular potassium". Neurochemistry International. 36 (4–5): 291–300. doi:10.1016/S0197-0186(99)00137-0. PMID 10732996.
Gabriel S, Njunting M, Pomper JK, Merschhemke M, Sanabria ER, Eilers A, Kivi A, Zeller M, Meencke HJ, Cavalheiro EA, Heinemann U, Lehmann TN (November 2004). "Stimulus and potassium-induced epileptiform activity in the human 齿状回 from 病人 with and without hippocampal sclerosis". The Journal of Neuroscience. 24 (46): 10416–30. doi:10.1523/JNEUROSCI.2074-04.2004. PMID 15548657.
Piet R, Vargová L, Syková E, Poulain DA, Oliet SH (February 2004). "Physiological contribution of the astrocytic environment of neurons to intersynaptic crosstalk". Proceedings of the National Academy of Sciences of the United States of America. 101 (7): 2151–5. Bibcode:2004PNAS..101.2151P. doi:10.1073/pnas.0308408100. PMC 357067. PMID 14766975.
Pascual O, Casper KB, Kubera C, Zhang J, Revilla-Sanchez R, Sul JY, Takano H, Moss SJ, McCarthy K, Haydon PG (October 2005). "Astrocytic purinergic signaling coordinates synaptic networks". Science. 310 (5745): 113–6. Bibcode:2005Sci...310..113P. doi:10.1126/science.1116916. PMID 16210541.
Parri R, Crunelli V (January 2003). "An astrocyte bridge from synapse to blood flow". Nature Neuroscience. 6 (1): 5–6. doi:10.1038/nn0103-5. PMID 12494240.
Ishibashi T, Dakin KA, Stevens B, Lee PR, Kozlov SV, Stewart CL, Fields RD (March 2006). "Astrocytes promote 髓磷脂ation in response to electrical impulses". Neuron. 49 (6): 823–32. doi:10.1016/j.neuron.2006.02.006. PMC 1474838. PMID 16543131.
Anderson MA, Burda JE, Ren Y, Ao Y, O'Shea TM, Kawaguchi R, Coppola G, Khakh BS, Deming TJ, Sofroniew MV (April 2016). "Astrocyte scar formation aids central nervous system axon regeneration". Nature. 532 (7598): 195–200. Bibcode:2016Natur.532..195A. doi:10.1038/nature17623. PMC 5243141. PMID 27027288.
Liddelow SA, Guttenplan KA, Clarke LE, Bennett FC, Bohlen CJ, Schirmer L, Bennett ML, Münch AE, Chung WS, Peterson TC, Wilton DK, Frouin A, Napier BA, Panicker N, Kumar M, Buckwalter MS, Rowitch DH, Dawson VL, Dawson TM, Stevens B, Barres BA (January 2017). "Neurotoxic reactive astrocytes are induced by activated microglia". Nature. 541 (7638): 481–487. Bibcode:2017Natur.541..481L. doi:10.1038/nature21029. PMC 5404890. PMID 28099414.
Han X, Chen M, Wang F, Windrem M, Wang S, Shanz S, Xu Q, Oberheim NA, Bekar L, Betstadt S, Silva AJ, Takano T, Goldman SA, Nedergaard M (March 2013). "Forebrain engraftment by human glial progenitor cells enhances synaptic plasticity and learning in adult mice". Cell Stem Cell. 12 (3): 342–53. doi:10.1016/j.stem.2012.12.015. PMC 3700554. PMID 23472873.
Brancaccio M, Edwards MD, Patton AP, Smyllie NJ, Chesham JE, Maywood ES, Hastings MH (January 2019). "Cell-autonomous clock of astrocytes drives circadian behavior in mammals". Science. 363 (6423): 187–192. doi:10.1126/science.aat4104. PMC 6440650. PMID 30630934.
Nossenson N, Magal A, Messer H (2016). "Detection of stimuli from multi-neuron activity: Empirical study and theoretical implications". Neurocomputing. 174: 822–837. doi:10.1016/j.neucom.2015.10.007.
Nossenson N (2013). Model Based Detection of a Stimulus Presence from Neurophysiological Signals (PDF). The Neiman Library of Exact Sciences & Engineering, Tel Aviv University: PhD diss, University of Tel-Aviv.
Cornell-Bell AH, Finkbeiner SM, Cooper MS, Smith SJ (January 1990). "Glutamate induces calcium waves in cultured astrocytes: long-range glial signaling". Science. 247 (4941): 470–3. Bibcode:1990Sci...247..470C. doi:10.1126/science.1967852. PMID 1967852.
Jahromi BS, Robitaille R, Charlton MP (June 1992). "Transmitter release increases intracellular calcium in perisynaptic Schwann cells in situ". Neuron. 8 (6): 1069–77. doi:10.1016/0896-6273(92)90128-Z. PMID 1351731.
Verkhratsky A, Orkand RK, Kettenmann H (January 1998). "Glial calcium: homeostasis and signaling function". Physiological Reviews. 78 (1): 99–141. doi:10.1152/physrev.1998.78.1.99. PMID 9457170.
Ebert U, Koch M (September 1997). "Acoustic startle-evoked potentials in the rat amygdala: effect of kindling". Physiology & Behavior. 62 (3): 557–62. doi:10.1016/S0031-9384(97)00018-8. PMID 9272664.
Frot M, Magnin M, Mauguière F, Garcia-Larrea L (March 2007). "Human SII and posterior 脑岛 differently encode thermal laser stimuli". Cerebral Cortex. 17 (3): 610–20. doi:10.1093/cercor/bhk007. PMID 16614165.
Perlman, Ido. "The Electroretinogram: ERG by Ido Perlman – Webvision". webvision.med.utah.edu.
Tian GF, Azmi H, Takano T, Xu Q, Peng W, Lin J, Oberheim N, Lou N, Wang X, Zielke HR, Kang J, Nedergaard M (September 2005). "An astrocytic basis of epilepsy". Nature Medicine. 11 (9): 973–81. doi:10.1038/nm1277. PMC 1850946. PMID 16116433.
Hertz L, Schousboe A, Boechler N, Mukerji S, Fedoroff S (February 1978). "Kinetic characteristics of the glutamate uptake into normal astrocytes in cultures". Neurochemical Research. 3 (1): 1–14. doi:10.1007/BF00964356. PMID 683409.
Bennett MV, Contreras JE, Bukauskas FF, Sáez JC (November 2003). "New roles for astrocytes: gap junction hemichannels have something to communicate". Trends in Neurosciences. 26 (11): 610–7. doi:10.1016/j.tins.2003.09.008. PMC 3694339. PMID 14585601.
Newman EA (April 2001). "Propagation of intercellular calcium waves in retinal astrocytes and Müller cells". The Journal of Neuroscience. 21 (7): 2215–23. doi:10.1523/JNEUROSCI.21-07-02215.2001. PMC 2409971. PMID 11264297.
Parpura V, Haydon PG (July 2000). "Physiological astrocytic calcium levels stimulate glutamate release to modulate adjacent neurons". Proceedings of the National Academy of Sciences of the United States of America. 97 (15): 8629–34. Bibcode:2000PNAS...97.8629P. doi:10.1073/pnas.97.15.8629. PMC 26999. PMID 10900020.
López-Caamal F, Oyarzún DA, Middleton RH, García MR (May 2014). "Spatial Quantification of Cytosolic Ca2⁺ Accumulation in Nonexcitable Cells: An Analytical Study". IEEE/ACM Transactions on Computational Biology and Bioinformatics. 11 (3): 592–603. doi:10.1109/TCBB.2014.2316010. PMID 26356026.
Haydon PG (March 2001). "GLIA: listening and talking to the synapse" (PDF). Nature Reviews. Neuroscience. 2 (3): 185–93. doi:10.1038/35058528. PMID 11256079.
Astrocytomas Archived 2012-04-05 at the Wayback Machine. International RadioSurgery Association (2010).
Astrocytoma Tumors. American Association of Neurological Surgeons (August 2005).
Barker AJ, Ullian EM (2008). "New roles for astrocytes in 发展 synaptic circuits". Communicative & Integrative Biology. 1 (2): 207–11. doi:10.4161/cib.1.2.7284. PMC 2686024. PMID 19513261.
Sloan SA, Barres BA (August 2014). "Mechanisms of astrocyte development and their contributions to neurodevelopmental disorders". Current Opinion in Neurobiology. 27: 75–81. doi:10.1016/j.conb.2014.03.005. PMC 4433289. PMID 24694749.
Garrison CJ, Dougherty PM, Kajander KC, Carlton SM (November 1991). "Staining of glial fibrillary acidic protein (GFAP) in lumbar spinal cord increases following a sciatic nerve constriction injury". Brain Research. 565 (1): 1–7. doi:10.1016/0006-8993(91)91729-K. PMID 1723019.
Volterra A & Meldolesi J (August 2005). "Astrocytes, from brain glue to communication elements: the revolution continues". Nature Reviews. Neuroscience. 6 (8): 626–40. doi:10.1038/nrn1722. PMID 16025096.
Halassa MM, Fellin T & Haydon PG (February 2007). "The tripartite synapse: roles for gliotransmission in health and disease". Trends in Molecular Medicine. 13 (2): 54–63. doi:10.1016/j.molmed.2006.12.005. PMID 17207662.
Sofroniew MV (November 2014). "Astrogliosis". Cold Spring Harbor Perspectives in Biology. 7 (2): a020420. doi:10.1101/cshperspect.a020420. PMC 4315924. PMID 25380660.
Sllvander, Sofia; Nikitidou, Elisabeth; Brolin, Robin; Sderberg, Linda; Sehlin, Dag; Lannfelt, Lars; Erlandsson, Anna (12 May 2016). "Accumulation of amyloid-β by astrocytes result in enlarged endosomes and microvesicle-induced apoptosis of neurons". Molecular Neurodegeneration. 11 (1): 38. doi:10.1186/s13024-016-0098-z. ISSN 1750-1326. PMC 4865996. PMID 27176225.
Bhat, Rekha; Crowe, Elizabeth P.; Bitto, Alessandro; Moh, Michelle; Katsetos, Christos D.; Garcia, Fernando U.; Johnson, Frederick Bradley; Trojanowski, John Q.; Sell, Christian; Torres, Claudio (2012-09-12). "Astrocyte Senescence as a Component of Alzheimer's Disease". PLoS ONE. 7 (9): e45069. doi:10.1371/journal.pone.0045069. ISSN 1932-6203.
Rostami, Jinar; Holmqvist, Staffan; Lindstrm, Veronica; Sigvardson, Jessica; Westermark, Gunilla T.; Ingelsson, Martin; Bergstrm, Joakim; Roybon, Laurent; Erlandsson, Anna (6 December 2017). "Human Astrocytes Transfer Aggregated α-Synuclein via Tunneling Nanotubes". The Journal of Neuroscience. 37 (49): 11835–11853. doi:10.1523/JNEUROSCI.0983-17.2017. ISSN 1529-2401. PMC 5719970. PMID 29089438.
Maragakis, Nicholas J; Rothstein, Jeffrey D (December 2006). "Mechanisms of Disease: astrocytes in neurodegenerative disease". Nature Clinical Practice Neurology. 2 (12): 679–689. doi:10.1038/ncpneuro0355. ISSN 1745-834X.
Davies SJ, Shih CH, Noble M, Mayer-Proschel M, Davies JE, Proschel C (March 2011). Combs C (ed.). "Transplantation of specific human astrocytes promotes functional recovery after spinal cord injury". PLOS ONE. 6 (3): e17328. Bibcode:2011PLoSO...617328D. doi:10.1371/journal.pone.0017328. PMC 3047562. PMID 21407803.
Yang Y, Ge W, Chen Y, Zhang Z, Shen W, Wu C, Poo M, Duan S (December 2003). "Contribution of astrocytes to hippocampal long-term potentiation through release of D-serine". Proceedings of the National Academy of Sciences of the United States of America. 100 (25): 15194–9. Bibcode:2003PNAS..10015194Y. doi:10.1073/pnas.2431073100. PMC 299953. PMID 14638938.
Jiao JW, Feldheim DA, Chen DF (June 2008). "Ephrins as negative regulators of adult neurogenesis in diverse regions of the central nervous system". Proceedings of the National Academy of Sciences of the United States of America. 105 (25): 8778–83. Bibcode:2008PNAS..105.8778J. doi:10.1073/pnas.0708861105. PMC 2438395. PMID 18562299.
Krencik R, Weick JP, Liu Y, Zhang ZJ, Zhang SC (May 2011). "Specification of transplantable astroglial sub类型 from human pluripotent stem cells". Nature Biotechnology. 29 (6): 528–34. doi:10.1038/nbt.1877. PMC 3111840. PMID 21602806.. Lay summary: Human Astrocytes Cultivated From Stem Cells In Lab Dish by U of Wisconsin Researchers. sciencedebate.com (22 May 2011)
Han J, Kesner P, Metna-Laurent M, Duan T, Xu L, Georges F, Koehl M, Abrous DN, Mendizabal-Zubiaga J, Grandes P, Liu Q, Bai G, Wang W, Xiong L, Ren W, Marsicano G, Zhang X (March 2012). "Acute cannabinoids impair working memory through astroglial CB1 receptor modulation of hippocampal LTD". Cell. 148 (5): 1039–50. doi:10.1016/j.cell.2012.01.037. PMID 22385967.
Gregori N, Prschel C, Noble M, Mayer-Prschel M (January 2002). "The tripotential glial-restricted precursor (GRP) cell and glial development in the spinal cord: generation of bipotential 少突胶质细胞-type-2 astrocyte progenitor cells and dorsal-ventral differences in GRP cell function". The Journal of Neuroscience. 22 (1): 248–56. doi:10.1523/JNEUROSCI.22-01-00248.2002. PMID 11756508.
Fulton BP, Burne JF, Raff MC (December 1992). "Visualization of O-2A progenitor cells in 发展 and adult rat 视神经 by quisqualate-stimulated cobalt uptake". The Journal of Neuroscience. 12 (12): 4816–33. doi:10.1523/JNEUROSCI.12-12-04816.1992. PMID 1281496.
Levison SW, Goldman JE (February 1993). "Both 少突胶质细胞s and astrocytes develop from progenitors in the subventricular zone of postnatal rat forebrain". Neuron. 10 (2): 201–12. doi:10.1016/0896-6273(93)90311-E. PMID 8439409.
Zerlin M, Levison SW, Goldman JE (November 1995). "Early patterns of migration, morphogenesis, and intermediate filament expression of subventricular zone cells in the postnatal rat forebrain". The Journal of Neuroscience. 15 (11): 7238–49. doi:10.1523/JNEUROSCI.15-11-07238.1995. PMID 7472478.
Brawer JR, Stein R, Small L, Cissé S, Schipper HM (November 1994). "Composition of Gomori-positive inclusions in astrocytes of the hypothalamic arcuate nucleus". The Anatomical Record. 240 (3): 407–15. doi:10.1002/ar.1092400313. PMID 7825737.
Young JK, McKenzie JC (November 2004). "GLUT2 immunoreactivity in Gomori-positive astrocytes of the hypo丘脑". The Journal of Histochemistry and Cytochemistry. 52 (11): 1519–24. doi:10.1369/jhc.4A6375.2004. PMC 3957823. PMID 15505347.
Marty N, Dallaporta M, Foretz M, Emery M, Tarussio D, Bady I, Binnert C, Beermann F, Thorens B (December 2005). "Regulation of glucagon secretion by glucose transporter type 2 (glut2) and astrocyte-dependent glucose sensors". The Journal of Clinical Investigation. 115 (12): 3545–53. doi:10.1172/jci26309. PMC 1297256. PMID 16322792.
Choi BH, Lapham LW (June 1978). "桡骨 glia in the human fetal cerebrum: a combined Golgi, immunofluorescent and electron microscopic study". Brain Research. 148 (2): 295–311. doi:10.1016/0006-8993(78)90721-7. PMID 77708.
Schmechel DE, Rakic P (June 1979). "A Golgi study of 桡骨 glial cells in 发展 monkey telencephalon: morphogenesis and transformation into astrocytes". Anatomy and Embryology. 156 (2): 115–52. doi:10.1007/BF00300010. PMID 111580.
Misson JP, Edwards MA, Yamamoto M, Caviness VS (November 1988). "Identification of 桡骨 glial cells within the 发展 murine central nervous system: studies based upon a new immunohistochemical marker". Brain Research. Developmental Brain Research. 44 (1): 95–108. doi:10.1016/0165-3806(88)90121-6. PMID 3069243.
Voigt T (November 1989). "Development of glial cells in the cerebral wall of ferrets: direct tracing of their transformation from 桡骨 glia into astrocytes". The Journal of Comparative Neurology. 289 (1): 74–88. doi:10.1002/cne.902890106. PMID 2808761.
Goldman SA, Zukhar A, Barami K, Mikawa T, Niedzwiecki D (August 1996). "室管膜l/sub室管膜l zone cells of postnatal and adult songbird brain generate both neurons and nonneuronal siblings in vitro and in vivo". Journal of Neurobiology. 30 (4): 505–20. doi:10.1002/(SICI)1097-4695(199608)30:4<505::AID-NEU6>3.0.CO;2-7. PMID 8844514.
Cisneros IE, Ghorpade A (October 2014). "Methamphetamine and HIV-1-induced neurotoxicity: role of trace amine associated receptor 1 cAMP signaling in astrocytes". Neuropharmacology. 85: 499–507. doi:10.1016/j.neuropharm.2014.06.011. PMC 4315503. PMID 24950453. Moreover, TAAR1 overexpression significantly decreased EAAT-2 levels and glutamate clearance that were further reduced by METH. Taken together, our data show that METH treatment activated TAAR1 leading to intracellular cAMP in human astrocytes and modulated glutamate clearance abilities. Furthermore, molecular alterations in astrocyte TAAR1 levels correspond to changes in astrocyte EAAT-2 levels and function. |

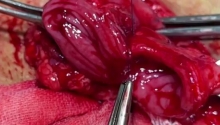
 因缺血而行小肠分段切除及一期吻合术
因缺血而行小肠分段切除及一期吻合术
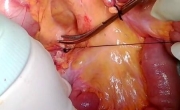
 肌肉活检 _ 如何进行肌肉活检
肌肉活检 _ 如何进行肌肉活检
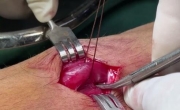
 尿道下裂修复后包皮瘘和龟头隐裂
尿道下裂修复后包皮瘘和龟头隐裂
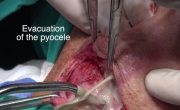 通过外部方法对喉囊肿进行有袋化或去顶
通过外部方法对喉囊肿进行有袋化或去顶

 发表于 2019-12-18 00:00:15
发表于 2019-12-18 00:00:15
 提升卡
提升卡 置顶卡
置顶卡 沉默卡
沉默卡 喧嚣卡
喧嚣卡 变色卡
变色卡 千斤顶
千斤顶 显身卡
显身卡