马上注册,结交更多好友,享用更多功能,让你轻松玩转社区。
您需要 登录 才可以下载或查看,没有账号?注册
×
髓磷脂是一种富含脂质的(脂肪)物质,围绕着神经细胞轴突(神经系统的电线),以脑岛的方式将它们隔开,并提高了信息(编码为电脉冲)沿轴突传递的速率。髓磷脂化的轴突可以比作带有脑岛刺刺材料(髓磷脂)的电线(轴突)。但是,与电线上的塑料覆盖物不同,髓磷脂不会在轴突的整个长度上形成单个长鞘。而是,每个轴突鞘状脑岛在单个长段上都分布着轴突,并且通常,每个轴突都包含多个长的髓磷脂化段,这些段被称为Ranvier节点的短间隙彼此隔开。髓磷脂由称为少突胶质细胞的神经胶质细胞在中枢神经系统(CNS)中形成,而由称为许旺细胞的神经胶质细胞在外周神经系统(PNS)中形成。在中枢神经系统中,轴突将电编码的信息从一个神经细胞体携带到另一个。在PNS中,轴突将信息传递到肌肉和腺体,或从皮肤等感觉器官传递信息。每个髓磷脂鞘是由少突胶质细胞(CNS)或雪旺细胞(PNS)过程(从细胞体到肢体的延伸)同心包裹在轴突周围形成的[1] [2]。
更具体地讲,髓磷脂通过脑岛固定轴突并减小轴突膜电容,从而加速了被称为动作电位[3]的电脉冲沿着沿脊髓标化的轴突的传递。在分子水平上,它增加了轴突外部的阳离子与在动作电位期间进入Ranvier节点处进入轴突并穿过髓鞘下方的轴突的Na离子之间的距离。这极大地减小了它们之间的排斥力的大小(根据库仑定律,排斥力与距离的平方成反比),否则它们将起到抑制钠离子运动的作用。
盐皮质传导中盐皮质鞘结构的不连续结构[4] [5] [6],其中动作电位从Ranvier的一个结点“跳越”到称为“节间”的轴突的较长的呈锥状的轴突上,然后“在Ranvier的下一个节点处重新充电”,依此类推,直到到达轴突终端为止。 Ranvier的节是相邻长(约0.2 mm –> 1 mm)呈螺旋状的节间之间的轴突的短(约1微米)非锥状区域。一旦到达轴突末端,该电信号就会激发一种化学信息或神经递质的释放,该信息或神经递质会与邻近突触后细胞(例如CNS中的神经细胞或PNS中的肌肉细胞)上的受体(称为突触)结合。
髓磷脂的这种“绝缘”作用对于正常的运动功能(例如,步行等运动),感觉功能(例如,听,见或感觉到疼痛的感觉)和认知(例如,获取和回忆知识)至关重要, 影响它的疾病的后果,例如遗传决定的白细胞营养不良; [7]后天性炎症性脱皮磷脂化疾病,多发性硬化症[8]和炎症性脱皮磷脂化周围神经病。[9] 由于多发性硬化症的患病率高,因此它是最著名的脊髓磷脂疾病,特别是影响中枢神经系统(大脑,脊髓和视神经)的多发性硬化症。

PNS中简化神经元的结构

CNS中具有少突胶质细胞和髓磷脂鞘的神经元
内容
1 发展
2 分布
3 组成
4 功能
5 临床意义
5.1 脱磷脂
5.1.1 症状
5.1.2 髓磷脂修复
5.2 脂质磷脂化
6 无脊椎动物髓磷脂
7 参考
发展历程
产生髓磷脂的过程称为髓磷脂形成或髓磷脂生成。在中枢神经系统中,称为少突胶质细胞前体细胞(OPC;少突胶质细胞的前体)的细胞分化为成熟的少突胶质细胞前体,形成髓磷脂。在人类中,虽然在出生时CNS或PNS中只有很少的髓磷脂存在,但在其妊娠的三个月中早期才开始发生髓磷脂。在婴儿期,随着人们越来越多的轴突获得了髓鞘,髓磷脂的发展迅速。这与认知和运动技能的发展相对应,包括语言理解,语音获取,爬行和行走。整个青春期和成年初期都会持续发生皮质磷脂化,尽管此时大部分已完成,但终生仍可在诸如大脑皮层等灰质区域添加髓磷脂鞘[11] [12] [13]。
物种分布
尽管轴突被无脊椎动物的一种称为神经胶质细胞的细胞所包裹,但它被认为是有颌脊椎动物(咬足动物)的定义特征。[14] [15]这些神经胶质膜包裹物与脊椎动物的紧凑型髓磷脂完全不同,如上所述,其通过围绕轴突多次同心包裹髓磷脂细胞过程而形成。 Rudolf Virchow于1854年首次描述了髓磷脂,尽管一个多世纪之后,随着电子显微镜的发展,其神经胶质细胞的起源和超微结构才变得显而易见。[17]
在脊椎动物中,并非所有的轴突都被髓磷脂化。例如,在PNS中,很大一部分轴突未被胶化。取而代之的是,它们被称为Remak SC的非髓鞘状Schwann细胞包裹,并排列成Remak束。[18]在中枢神经系统中,非髓磷脂样的轴突(或间歇性的髓磷脂样轴突,指的是在髓磷脂样段之间有较长的非髓磷脂样区域的轴突)与髓磷脂样的轴突交织在一起,并至少部分地被交织在一起。另一种称为星形胶质细胞的神经胶质细胞的过程。
组成

康涅狄格州哈特福德市三一学院的电子显微镜工厂产生的锥状PNS轴突横截面的透射电子显微照片

锥状轴突的横截面图
轴突
雪旺细胞核
雪旺氏细胞
髓磷脂鞘
神经质
CNS髓磷脂的成分和构型与PNS髓磷脂略有不同,但两者都具有相同的“脑岛刺”功能(见上文)。脂质磷脂富含脂质,因此呈白色,因此被称为CNS的“白质”。中枢神经系统白质束(例如视神经,皮质脊髓束和)体)和PNS神经(例如坐骨神经和听觉神经,也呈现白色)均包含成千上万的轴突,大部分平行排列。血管为氧气和能量底物(例如葡萄糖)提供了到达这些纤维束的途径,这些纤维束还包含其他细胞类型,包括CNS中的星形胶质细胞和小胶质细胞以及PNS中的巨噬细胞。
就总质量而言,髓磷脂包含约40%的水;干物质包含60%至75%的脂质和15%至25%的蛋白质。蛋白质含量包括髓磷脂碱性蛋白(MBP),[19]在中枢神经系统中含量丰富,在形成致密的髓磷脂中起着至关重要的,非冗余的作用。皮质磷脂少突胶质细胞糖蛋白(MOG),[20]对CNS具有特异性;蛋白脂蛋白(PLP),[21]是中枢神经系统髓磷脂中含量最丰富的蛋白,但仅是PNS髓磷脂的一小部分。在PNS中,髓磷脂零蛋白(MPZ或P0)与CNS中的PLP具有相似的作用,因为它参与将构成髓磷脂鞘的神经胶质细胞膜的多个同心层保持在一起。骨髓磷脂的主要脂质是称为半乳糖脑苷的糖脂。 sphingo锥磷脂的缠绕的碳氢链增强了髓磷脂鞘。胆固醇是髓磷脂的必不可少的脂质成分,没有胆固醇脂质就无法形成。[22]
功能
主条目:盐度传导

由于盐碱化传导,动作神经元在髓磷脂样神经元中的传播速度要快于非髓磷脂样神经元。
锥形磷脂的主要目的是提高电脉冲沿着锥形磷脂纤维传播的速度。在非髓磷脂纤维中,电脉冲(动作电位)以连续波的形式传播,但在髓磷脂纤维中,它们通过跳跃传导“跳跃”或传播。至少对于一定直径的轴突,后者明显比前者快。髓磷脂降低了电容,并增加了整个轴突膜(axolemma)的电阻。有人提出,通过维持远端身体各部位之间的敏捷沟通,髓磷脂可以允许更大的体型。[14]
沿桡方向分布的纤维状纤维在沿轴向的节点间缺乏电压门控的钠通道,仅在Ranvier的节点处暴露。在这里,它们非常丰富且密密麻麻。[23]带正电的钠离子可以通过这些电压门控通道进入轴突,导致Ranvier节点处的膜电位去极化。然后,由于带正电的钾离子通过钾通道离开轴突,使静息膜电位迅速恢复。轴突内部的钠离子然后迅速扩散通过轴质(轴突细胞质),到达相邻的脊髓标间节点,最终到达Ranvier的下一个(远端)节点,触发电压门控钠通道的开放和钠离子的进入在这个网站上。尽管钠离子迅速扩散穿过腋质,但扩散本质上是递减的,因此Ranvier的结点必须(相对)紧密隔开,以确保动作电位的传播。[24]当腋静脉膜电位去极化至大约+35 mV时,动作电位在Ranvier的连续节点处“充电”。[23]沿锥磷脂化的节间,依赖于能量的钠/钾泵将钠离子从轴突中抽出,将钾离子泵回轴突中,以恢复细胞内(细胞内部,在这种情况下为轴突)之间的离子平衡。和细胞外(细胞外)液体。
众所周知,髓磷脂作为“轴突绝缘体”的作用是鲜为人知的,或者只是最近才确立。髓磷脂化细胞通过促进神经丝的磷酸化来“雕刻”下面的轴突,从而增加了结间区域的轴突的直径或厚度。帮助分子在Ranvier的节点上聚集在轴突上(例如电压门控钠通道); [25]并调节细胞骨架结构和细胞器(例如线粒体)沿轴突的运输。[26]在2012年,有证据显示支持髓磷脂细胞在“喂食”轴突中发挥作用。[27] [28]换句话说,髓磷脂细胞似乎是轴突的局部“加油站”,它使用大量能量来恢复轴突与其周围环境之间离子的正常平衡,[29] [30]动作电位的产生。
当外围纤维被切断时,髓磷脂鞘提供了一条眶,沿着该眶可发生再生长。但是,髓磷脂层不能确保神经纤维的完美再生。一些再生的神经纤维找不到正确的肌肉纤维,而周围神经系统的一些受损运动神经元则死而无再生长。脊髓磷脂鞘和神经纤维的损伤通常与功能不全增加有关。
哺乳动物中枢神经系统的非髓磷脂化的纤维和髓磷脂化的轴突不会再生。
一些研究表明,视神经纤维可以在出生后的大鼠中再生。这种再生取决于两个条件:必须用适当的神经营养因子防止轴突消亡,并且必须使神经突生长抑制成分失活。这些研究可能会导致对中枢神经系统中神经纤维再生的进一步了解。
临床意义
脱磷脂
更多信息:去除磷脂疾病
髓鞘磷脂化是神经系统中髓磷脂鞘脑岛的丧失,是一些神经退行性自身免疫疾病的标志,包括多发性硬化症,急性弥漫性脑脊髓炎,视神经脊髓炎,横贯性脊髓炎,慢性炎症性脱髓鞘性多发性神经病,格林-巴利综合征(Guillain-Barrésyndrome),中枢桥脑髓样磷脂病,遗传性髓鞘变性疾病(如白细胞营养不良和夏科特-玛丽-牙齿疾病)。如果不迅速诊断出这种病,恶性贫血的患者也会遭受神经损伤。继发于恶性贫血的亚急性合并脊髓变性可导致轻微的周围神经损害,严重损害中枢神经系统,影响言语,平衡和认知意识。当髓磷脂降解时,沿神经的信号传导会受到损害或丧失,神经最终会萎缩。[需要澄清]髓磷脂恶化的一种更严重的情况称为Canavan病。
免疫系统可能在与此类疾病相关的脱皮磷脂中发挥作用,包括炎症通过肿瘤坏死因子[31]或干扰素的上调引起的细胞因子的过量产生而导致脱皮磷脂化。
病征
脱髓磷脂结果导致多种症状,这些症状取决于受影响的神经元的功能。它破坏大脑和身体其他部位之间的信号;症状因患者而异,并且在临床观察和实验室研究中都有不同的表现。
典型症状包括仅影响一只眼睛的中央视野模糊,可能伴有眼球运动疼痛,复视,视力丧失/听力,腿,手臂,胸部或面部的奇怪感觉,例如刺痛或麻木。 (神经病),手臂或腿部无力,认知障碍,包括言语障碍和记忆力减退,热敏感性(在热淋淋下出现症状加重或再次出现症状,例如洗热水澡),敏捷度下降,协调动作困难或平衡障碍,难以控制排便或排尿,疲劳和耳鸣。[32]
髓磷脂修复
更多信息:视网膜磷脂
修复受损的血管内皮的研究正在进行中。技术包括通过外科手术将少突胶质细胞前体细胞植入中枢神经系统,并用某些抗体诱导髓磷脂修复。尽管小鼠的结果令人鼓舞(通过干细胞移植),但该技术是否能有效替代人的血浆磷脂损失尚不清楚。[33]胆碱能治疗,例如乙酰胆碱酯酶抑制剂(AChEIs),可能对髓鞘磷脂化,髓磷脂修复和髓磷脂完整性具有有益作用。胆碱能刺激的增加也可能通过对大脑发育过程,特别是对少突胶质细胞及其支持的终生血浆磷脂化过程的微妙营养作用起作用。增加少突胶质细胞的胆碱能刺激,AChEIs和其他胆碱能治疗,例如尼古丁,可能会促进发育过程中的髓磷脂形成和老年人的髓磷脂修复。[34]糖原合酶激酶3β抑制剂(如氯化锂)已被发现能促进面神经受损小鼠的髓磷脂形成。[35]胆固醇是维生素B12的必需品,是鞘磷脂的必需营养素。[36] [37]
脱髓鞘
脱髓鞘的特征在于髓磷脂鞘的结构和功能有缺陷。不像去血管磷脂,它不会产生病变。这种有缺陷的鞘层通常是由基因突变引起的,这些突变影响了髓磷脂的生物合成和形成。颤抖的小鼠代表了一种dys髓磷脂的动物模型。牵涉到神经纤维化异常的人类疾病包括白细胞营养不良(Pelizaeus–Merzbacher病,Canavan病,苯丙酮尿症)和精神分裂症。[38] [39] [40]
无脊椎动物髓磷脂
在几种无脊椎动物类群中发现了功能上等效的类磷脂样鞘,包括寡毛类,对虾类和类颅类。这些类似于鞘磷脂的鞘与脊椎动物的鞘具有一些结构特征,包括膜的多样性,膜的凝结和结节。[14]但是,脊椎动物的结点是环状的。即它们包围着轴突。相反,在无脊椎动物的鞘中发现的结节是环状的或有孔的。即它们仅限于“地点”。值得注意的是,最快的记录的传导速度(在脊椎动物和无脊椎动物中)在无脊椎动物虾的被包裹的轴突中发现,无脊椎动物[14]的范围为90到200 m / s [15](参见100–120)。 m / s表示最快的锥状脊椎动物轴突)。
另见
Lesional de髓磷脂ations of the central nervous system
髓磷脂 incisure
The 髓磷脂 Project, project to regenerate 髓磷脂
髓磷脂 修复 Foundation, a nonprofit medical research foundation for multiple sclerosis drug discovery.
参考
Stassart, Ruth M.; Mbius, Wiebke; Nave, Klaus-Armin; Edgar, Julia M. (2018). "The Axon-髓磷脂 Unit in Development and Degenerative Disease". Frontiers in Neuroscience. 12: 467. doi:10.3389/fnins.2018.00467. ISSN 1662-4548. PMC 6050401. PMID 30050403.
Stadelmann, Christine; Timmler, Sebastian; Barrantes-Freer, Alonso; Simons, Mikael (2019-07-01). "髓磷脂 in the Central Nervous System: 结构, Function, and Pathology". Physiological Reviews. 99 (3): 1381–1431. doi:10.1152/physrev.00031.2018. ISSN 1522-1210. PMID 31066630.
Bean, Bruce P. (June 2007). "The action potential in mammalian central neurons". Nature Reviews Neuroscience. 8 (6): 451–465. doi:10.1038/nrn2148. ISSN 1471-0048.
Carroll, SL. "The Molecular and Morphologic 结构s That Make Saltatory Conduction Possible in Peripheral Nerve". PMID 28340093.
Keizer J, Smith GD, Ponce-Dawson S, Pearson JE (August 1998). "Saltatory propagation of Ca2+ waves by Ca2+ sparks". Biophysical Journal. 75 (2): 595–600. Bibcode:1998BpJ....75..595K. doi:10.1016/S0006-3495(98)77550-2. PMC 1299735. PMID 9675162.
Dawson SP, Keizer J, Pearson JE (May 1999). "Fire-diffuse-fire model of dynamics of intracellular calcium waves". Proceedings of the National Academy of Sciences of the United States of America. 96 (11): 6060–3. Bibcode:1999PNAS...96.6060D. doi:10.1073/pnas.96.11.6060. PMC 26835. PMID 10339541.
van der Knaap MS, Bugiani M (September 2017). "Leukodystrophies: a proposed classification system based on pathological changes and pathogenetic mechanisms". Acta Neuropathologica. 134 (3): 351–382. doi:10.1007/s00401-017-1739-1. PMC 5563342. PMID 28638987.
Compston A, Coles A (October 2008). "Multiple sclerosis". Lancet. 372 (9648): 1502–17. doi:10.1016/S0140-6736(08)61620-7. PMID 18970977.
Lewis RA (October 2017). "Chronic inflammatory de髓磷脂ating polyneuropathy". Current Opinion in Neurology. 30 (5): 508–512. doi:10.1097/WCO.0000000000000481. PMID 28763304.
"Pediatric Neurologic Examination Videos & Descriptions: Developmental Anatomy". library.med.utah.edu. Retrieved 2016-08-20.
Swire M, Ffrench-Constant C (May 2018). "Seeing Is Believing: 髓磷脂 Dynamics in the Adult CNS". Neuron. 98 (4): 684–686. doi:10.1016/j.neuron.2018.05.005. PMID 29772200.
Hill RA, Li AM, Grutzendler J (May 2018). "Lifelong cortical 髓磷脂 plasticity and age-related degeneration in the live mammalian brain". Nature Neuroscience. 21 (5): 683–695. doi:10.1038/s41593-018-0120-6. PMC 5920745. PMID 29556031.
Hughes EG, Orthmann-Murphy JL, Langseth AJ, Bergles DE (May 2018). "髓磷脂 remodeling through experience-dependent oligodendrogenesis in the adult somatosensory cortex". Nature Neuroscience. 21 (5): 696–706. doi:10.1038/s41593-018-0121-5. PMC 5920726. PMID 29556025.
Hartline DK (May 2008). "What is 髓磷脂?". Neuron Glia Biology. 4 (2): 153–63. doi:10.1017/S1740925X09990263. PMID 19737435.
Salzer JL, Zalc B (October 2016). "髓磷脂ation" (PDF). Current Biology. 26 (20): R971–R975. Bibcode:1996CBio....6.1213A. doi:10.1016/j.cub.2016.07.074. PMID 27780071.
Virchow R (1854). "Ueber das ausgebreitete Vorkommen einer dem Nervenmark analogen Substanz in den thierischen Geweben". Archiv für Pathologische Anatomie und Physiologie und für Klinische Medizin (in German). 6 (4): 562–572. doi:10.1007/BF02116709.
Boullerne AI (September 2016). "The history of 髓磷脂". Experimental Neurology. 283 (Pt B): 431–445. doi:10.1016/j.expneurol.2016.06.005. PMC 5010938. PMID 27288241.
Monk KR, Feltri ML, Taveggia C (August 2015). "New insights on Schwann cell development". Glia. 63 (8): 1376–1393. doi:10.1002/glia.22852. PMC 4470834. PMID 25921593.
Steinman L (May 1996). "Multiple sclerosis: a coordinated immunological attack against 髓磷脂 in the central nervous system". Cell. 85 (3): 299–302. doi:10.1016/S0092-8674(00)81107-1. PMID 8616884.
Mallucci G, Peruzzotti-Jametti L, Bernstock JD, Pluchino S (April 2015). "The role of immune cells, glia and neurons in white and gray matter pathology in multiple sclerosis". Progress in Neurobiology. 127-128: 1–22. doi:10.1016/j.pneurobio.2015.02.003. PMC 4578232. PMID 25802011.
Greer JM, Lees MB (March 2002). "髓磷脂 proteolipid protein--the first 50 years". The International Journal of Biochemistry & Cell Biology. 34 (3): 211–215. doi:10.1016/S1357-2725(01)00136-4. PMID 11849988.
Saher G, Brügger B, Lappe-Siefke C, Mbius W, Tozawa R, Wehr MC, Wieland F, Ishibashi S, Nave KA (April 2005). "High cholesterol level is essential for 髓磷脂 membrane growth". Nature Neuroscience. 8 (4): 468–475. doi:10.1038/nn1426. PMID 15793579.
Saladin KS (2012). Anatomy & physiology: the unity of form and function (6th ed.). New York, NY: McGraw-Hill.[page needed]
Raine CS (1999). "Characteristics of Neuroglia". In Siegel GJ, Agranoff BW, Albers RW, Fisher SK, Uhler MD (eds.). Basic Neurochemistry: Molecular, Cellular and Medical Aspects (6th ed.). Philadelphia: Lippincott-Raven.
Brivio V, Faivre-Sarrailh C, Peles E, Sherman DL, Brophy PJ (April 2017). "Assembly of CNS Nodes of Ranvier in 髓磷脂ated Nerves Is Promoted by the Axon Cytoskeleton". Current Biology. 27 (7): 1068–1073. Bibcode:1996CBio....6.1213A. doi:10.1016/j.cub.2017.01.025. PMC 5387178. PMID 28318976.
Stassart RM, Mbius W, Nave KA, Edgar JM (2018). "The Axon-髓磷脂 Unit in Development and Degenerative Disease". Frontiers in Neuroscience. 12: 467. doi:10.3389/fnins.2018.00467. PMC 6050401. PMID 30050403.
Fünfschilling U, Supplie LM, Mahad D, Boretius S, Saab AS, Edgar J, Brinkmann BG, Kassmann CM, Tzvetanova ID, Mbius W, Diaz F, Meijer D, Suter U, Hamprecht B, Sereda MW, Moraes CT, Frahm J, Goebbels S, Nave KA (April 2012). "Glycolytic 少突胶质细胞s maintain 髓磷脂 and long-term axonal integrity". Nature. 485 (7399): 517–521. Bibcode:2012Natur.485..517F. doi:10.1038/nature11007. PMC 3613737. PMID 22622581.
Lee Y, Morrison BM, Li Y, Lengacher S, Farah MH, Hoffman PN, Liu Y, Tsingalia A, Jin L, Zhang PW, Pellerin L, Magistretti PJ, Rothstein JD (July 2012). "Oligodendroglia metabolically support axons and contribute to neurodegeneration". Nature. 487 (7408): 443–448. Bibcode:2012Natur.487..443L. doi:10.1038/nature11314. PMC 3408792. PMID 22801498.
Engl E, Attwell D (August 2015). "Non-signalling energy use in the brain". The Journal of Physiology. 593 (16): 3417–329. doi:10.1113/jphysiol.2014.282517. PMC 4560575. PMID 25639777.
Attwell D, Laughlin SB (October 2001). "An energy budget for signaling in the grey matter of the brain". Journal of Cerebral Blood Flow and Metabolism. 21 (10): 1133–145. doi:10.1097/00004647-200110000-00001. PMID 11598490.
Ledeen RW, Chakraborty G (March 1998). "Cytokines, signal transduction, and inflammatory de髓磷脂ation: review and hypothesis". Neurochemical Research. 23 (3): 277–289. doi:10.1023/A:1022493013904. PMID 9482240.
Mayo Clinic 2007 and University of Leicester Clinical Studies, 2014[full citation needed]
Windrem MS, Nunes MC, Rashbaum WK, Schwartz TH, Goodman RA, McKhann G, Roy NS, Goldman SA (January 2004). "Fetal and adult human 少突胶质细胞 progenitor cell isolates 髓磷脂ate the congenitally dys髓磷脂ated brain". Nature Medicine. 10 (1): 93–97. doi:10.1038/nm974. PMID 14702638. Lay summary – FuturePundit.
Bartzokis G (August 2007). "Acetylcholinesterase inhibitors may improve 髓磷脂 integrity". Biological Psychiatry. 62 (4): 294–301. doi:10.1016/j.biopsych.2006.08.020. PMID 17070782.
Makoukji J, Belle M, Meffre D, Stassart R, Grenier J, Shackleford G, Fledrich R, Fonte C, Branchu J, Goulard M, de Waele C, Charbonnier F, Sereda MW, Baulieu EE, Schumacher M, Bernard S, Massaad C (March 2012). "Lithium enhances re髓磷脂ation of peripheral nerves". Proceedings of the National Academy of Sciences of the United States of America. 109 (10): 3973–3978. Bibcode:2012PNAS..109.3973M. doi:10.1073/pnas.1121367109. PMC 3309729. PMID 22355115.
Petrov AM, Kasimov MR, Zefirov AL (2016). "Brain Cholesterol Metabolism and Its Defects: Linkage to Neurodegenerative Diseases and Synaptic Dysfunction". Acta Naturae. 8 (1): 58–73. doi:10.32607/20758251-2016-8-1-58-73. PMC 4837572. PMID 27099785.
Miller A, Korem M, Almog R, Galboiz Y (June 2005). "Vitamin B12, de髓磷脂ation, re髓磷脂ation and 修复 in multiple sclerosis". Journal of the Neurological Sciences. 233 (1–2): 93–97. doi:10.1016/j.jns.2005.03.009. PMID 15896807.
Krämer-Albers EM, Gehrig-Burger K, Thiele C, Trotter J, Nave KA (November 2006). "Perturbed interactions of mutant proteolipid protein/DM20 with cholesterol and lipid rafts in oligodendroglia: implications for dys髓磷脂ation in spastic paraplegia". The Journal of Neuroscience. 26 (45): 11743–1752. doi:10.1523/JNEUROSCI.3581-06.2006. PMID 17093095.
Matalon R, Michals-Matalon K, Surendran S, Tyring SK (2006). Canavan disease: studies on the knockout mouse. Advances in Experimental Medicine and Biology. 576. pp. 77–93, discussion 361–363. doi:10.1007/0-387-30172-0_6. ISBN 978-0-387-30171-6. PMID 16802706.
Tkachev D, Mimmack ML, Huffaker SJ, Ryan M, Bahn S (August 2007). "Further evidence for altered 髓磷脂 biosynthesis and glutamatergic dysfunction in schizophrenia". The International Journal of Neuropsychopharmacology. 10 (4): 557–563. doi:10.1017/S1461145706007334. PMID 17291371. |

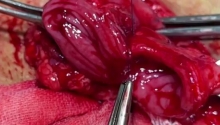
 因缺血而行小肠分段切除及一期吻合术
因缺血而行小肠分段切除及一期吻合术
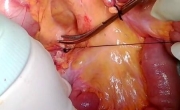
 肌肉活检 _ 如何进行肌肉活检
肌肉活检 _ 如何进行肌肉活检
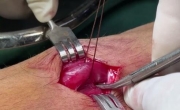
 尿道下裂修复后包皮瘘和龟头隐裂
尿道下裂修复后包皮瘘和龟头隐裂
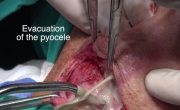 通过外部方法对喉囊肿进行有袋化或去顶
通过外部方法对喉囊肿进行有袋化或去顶

 发表于 2019-12-16 00:00:12
发表于 2019-12-16 00:00:12
 提升卡
提升卡 置顶卡
置顶卡 沉默卡
沉默卡 喧嚣卡
喧嚣卡 变色卡
变色卡 千斤顶
千斤顶 显身卡
显身卡