马上注册,结交更多好友,享用更多功能,让你轻松玩转社区。
您需要 登录 才可以下载或查看,没有账号?注册
×
颗粒细胞这个名称已被用于许多不同类型的神经元,它们的唯一共同特征是它们都具有非常小的细胞体。颗粒细胞位于小脑的颗粒层,海马的齿状回,背耳蜗核的表层,嗅球和大脑皮层内。
小脑颗粒细胞占人脑神经元的大多数。这些颗粒细胞接受源自脑桥核的苔藓纤维的兴奋性输入。小脑颗粒细胞通过Purkinje层向上突出到分子层,在其中分支成平行纤维,这些平行纤维通过浦肯野细胞树突状树突扩散。这些平行纤维使用谷氨酸作为神经递质,在浦肯野细胞的中间和远端树突上形成数千个兴奋性颗粒细胞-浦肯野细胞突触。
大脑皮层的第4层颗粒细胞从丘脑接收输入,并将投影发送到大脑皮层的2-3层,也发送到大脑皮层的颗粒下的层。
 and granule cells (B) from pigeon 小脑 by Sa.jpg")
1899年,圣地亚哥·拉蒙·卡哈尔(SantiagoRamóny Cajal)从小脑中抽出浦肯野细胞(A)和颗粒细胞(B)。西班牙马德里圣地亚哥·拉蒙·卡哈尔研究所。
内容
1 结构
1.1 小脑颗粒细胞
1.2 齿状回颗粒细胞
1.3 背耳蜗核颗粒细胞
1.4 嗅球颗粒细胞
2 功能
2.1 小脑的神经通路和回路
2.2 小脑颗粒细胞上的苔藓纤维输入
2.3 在小脑颗粒细胞上爬纤维输入
2.4 不同颗粒细胞的特殊功能
2.5 功能的关键因素
3 在疾病中的作用
3.1 齿状颗粒细胞的形态改变
3.2 颗粒细胞神经发生减少
3.3 较短的颗粒细胞树突
4 参考
结构
不同大脑区域的颗粒细胞在功能和解剖学上都不同:它们唯一的共同点是体积小。例如,嗅球颗粒细胞具有GABA能和无轴突作用,而齿状回中的颗粒细胞具有谷氨酸能投射轴突。这两个颗粒细胞群体也是经历成年神经发生的仅有的主要神经元群体,而小脑和皮质颗粒细胞则没有。颗粒细胞(除嗅球细胞外)具有神经元的典型结构,该神经元由树突,躯体(细胞体)和轴突组成。
树突:每个颗粒细胞都有3-4个粗大的树突,其末端为爪状。每个树突的长度仅约15μm。
体细胞:颗粒细胞均具有约10μm的较小体细胞直径。
轴突:每个颗粒细胞将一个轴突发送到浦肯野细胞树突状树上。轴突的直径极窄:微米。
突触:将100-300,000粒细胞轴突突触到单个浦肯野细胞上。
颗粒细胞之间的间隙连接的存在允许多个神经元彼此耦合,从而允许多个细胞同步发挥作用,并允许发生颗粒细胞发育所必需的信号传导功能。[1]
小脑颗粒细胞
主条目:小脑颗粒细胞
由菱形唇产生的颗粒细胞位于小脑皮质的颗粒细胞层中。它们很小,很多。它们的特征是非常小的躯体和几个短的树突,它们以爪形末端终止。在透射电子显微镜中,这些细胞的特征是细胞核边缘被深色染色的核染成深色。轴突上升到分子层,在那里分裂形成平行纤维。[2]
齿状回颗粒细胞
齿状回的主要细胞类型是颗粒细胞。齿状回颗粒细胞具有椭圆形的细胞体,其宽度约为10μm,高度约为18μm。[3]
颗粒细胞具有特征性的圆锥形的尖顶树突状树。树突分支遍布整个分子层,树突树的最尖端终止于海马裂隙或心室表面。[4]颗粒细胞紧密地堆积在齿状回的颗粒细胞层中。
背耳蜗核颗粒细胞
耳蜗背核中的颗粒细胞是具有两个或三个短树突的小神经元,在末端形成一些分支,并具有扩张。树突短,具爪状末端,形成小球以接收苔藓纤维,类似于小脑颗粒细胞。[5]它的轴突伸到耳蜗背核的分子层,在那里形成平行纤维,也类似于小脑颗粒细胞。[6]背耳蜗颗粒细胞是小的兴奋性中间神经元,与发育相关,因此类似于小脑颗粒细胞。
嗅球颗粒细胞
脊椎动物嗅球中的主要内在颗粒细胞缺乏轴突(副神经元也没有)。每个细胞产生短的中央树突和单个长的顶端树突,其扩展到颗粒细胞层并进入二瓣细胞体层。树突分支终止于嗅束中树突中的外部丛状层内。[7]在哺乳动物的嗅球中,由于存在大刺,颗粒细胞可以处理突触的输入和输出。[8]
功能

小脑的神经通路和回路
对小脑皮层中不同类型神经元之间存在的神经联系进行卡通重新介绍。包括浦肯野细胞,颗粒细胞和中间神经元。
小脑中的神经通路和回路
小脑颗粒细胞接受来自脑桥核的3或4苔藓纤维的兴奋性输入。苔藓纤维与颗粒细胞形成兴奋性连接,导致颗粒细胞激发动作电位。
小脑颗粒细胞的轴突裂开,形成支配浦肯野细胞的平行纤维。在平行纤维上发现了绝大多数的颗粒细胞轴突突触。[9]
平行纤维通过Purkinje层向上传送到分子层,在那里它们分支并通过浦肯野细胞树突状乔木扩散。这些平行纤维在浦肯野细胞的树突上形成成千上万的兴奋性颗粒细胞-浦肯野细胞突触。
随着谷氨酸的释放,这种连接是兴奋的。
来自同一颗粒细胞的平行纤维和上升轴突突触同步产生兴奋信号。小脑皮质中存在多种抑制性神经元。小脑皮层中唯一的兴奋性神经元是颗粒细胞。[10]
人们认为,平行纤维和浦肯野细胞之间突触的可塑性对于运动学习很重要。[11]小脑回路的功能完全取决于颗粒层执行的过程。因此,颗粒细胞的功能决定了整个小脑的功能。[12]
小脑颗粒细胞上的苔藓纤维输入
颗粒细胞的树突也与独特的非髓样轴突突触,SantiagoRamóny Cajal将其称为苔藓纤维[4]。苔藓纤维和高尔基体细胞均与颗粒细胞形成突触连接。这些细胞一起形成肾小球。[10]
颗粒细胞受到前馈抑制:颗粒细胞激发浦肯野细胞,但也会激发抑制浦肯野细胞的GABA能中间神经元。
颗粒细胞也受到反馈抑制:高尔基细胞受到颗粒细胞的兴奋性刺激,进而向颗粒细胞发送抑制信号。[13]
在颗粒细胞之间的突触传递过程中,苔藓纤维的输入代码得以保留,这表明神经支配特定于所接收的输入。[14]颗粒细胞不仅中继来自苔藓纤维的信号,而且还执行时空域所需的各种复杂转换。[10]
每个颗粒细胞都从两个不同的生苔细小输入接收输入。因此,输入来自两个不同的地方,而不是颗粒细胞从同一来源接收多个输入。
向颗粒细胞发送信号的苔藓纤维的差异直接影响颗粒细胞转化为浦肯野细胞的信息类型。这种转移的可靠性取决于颗粒细胞中突触活动的可靠性以及所接受刺激的性质。[15]颗粒细胞从苔藓纤维接收的信号取决于苔藓纤维本身的功能。因此,颗粒细胞能够整合来自不同苔藓纤维的信息并产生新的活动模式。[15]
在小脑颗粒细胞上爬入纤维
不同模式的长满苔藓的细密输入将在颗粒细胞中产生独特的活动模式,这些模式可以通过攀爬光纤输入所传达的示教信号进行修改。大卫·马尔(David Marr)和詹姆斯·阿布斯(James Albus)建议,小脑用作自适应滤波器,根据感觉输入的性质改变运动行为。
由于多个(〜200,000)颗粒细胞突触到单个浦肯野细胞上,因此可以响应来自攀爬纤维输入的“触发信号”来改变每个平行纤维的作用。
不同颗粒细胞的特定功能
小脑颗粒细胞
大卫·马尔(David Marr)提出,颗粒细胞编码长满苔藓的纤维的组合。为了使颗粒细胞做出反应,它需要接收来自多生苔纤维的有效输入。小脑中多个输入结果的组合比单个苔藓纤维所能允许的更精确地区分输入模式。[16]小脑颗粒细胞还与协调大脑中发现的GABA的周围水平一起,在控制睡眠的补品电导中起着协调作用。
齿状颗粒细胞
海马结果在空间记忆缺陷中丧失齿状回神经元。因此,齿状颗粒细胞被认为在空间记忆的形成中起作用。[17]未成熟和成熟的齿状颗粒细胞在记忆功能中具有不同的作用。成年颗粒细胞被认为与模式分离有关,而老颗粒细胞则有助于快速完成模式。[18]
背耳蜗颗粒细胞
来自初级听觉皮层的锥体细胞直接投射到耳蜗核上。这在听觉惊吓反射中很重要,在听觉惊吓反射中,脊髓细胞调节次级定向反射,而颗粒细胞的输入负责适当的定向。[19]这是因为颗粒细胞接收的信号包含有关头部位置的信息。耳蜗背核中的颗粒细胞在作者的环境中对声音的感知和响应中发挥作用。
嗅球颗粒细胞
由颗粒细胞产生的抑制作用是嗅球中最常见的GABA能细胞类型,在形成嗅球的输出中起着至关重要的作用。[20] GABA能颗粒细胞接受两种类型的兴奋性输入;被AMPA受体激活的那些和被NMDA受体激活的那些。这使颗粒细胞能够调节嗅球中感觉输入的过程。[20]嗅球将气味信息从鼻子传送到大脑,因此对于适当的嗅觉是必不可少的。人们还发现,嗅球中的颗粒细胞在形成与气味有关的记忆方面也很重要。[21]
功能的关键因素
钙
钙动力学对于颗粒细胞的多种功能至关重要,例如改变膜电位,突触可塑性,细胞凋亡和基因转录调控。[10]控制嗅球颗粒细胞棘突触前和突触后功能的钙信号的性质尚不清楚。[8]
一氧化氮
颗粒神经元具有高水平的一氧化氮合酶神经元同工型。该酶取决于钙的存在,并负责产生一氧化氮(NO)。该神经递质是颗粒细胞前体增殖的负调节剂,其促进不同颗粒细胞的分化。 NO调节颗粒细胞与神经胶质之间的相互作用[10],对于保护颗粒细胞免受损害至关重要。 NO还负责神经可塑性和运动学习。[22]
在疾病中的作用
齿状颗粒细胞的形态改变
TrkB负责维护齿状颗粒细胞的正常突触连接。 TrkB还调节颗粒细胞的特定形态(生物学),因此据称在调节神经元发育,神经元可塑性,学习和癫痫的发展中很重要。[23]粒细胞的TrkB调节对预防记忆缺陷和边缘性癫痫很重要。这是由于这样的事实,即齿状颗粒细胞在健康和疾病中在内脏海马回路的功能中起着关键作用。齿状颗粒细胞的位置可以调节信息向海马的流动,海马是正常学习和记忆所需的结构。[23]
颗粒细胞神经发生减少
癫痫和抑郁症均显示成年海马颗粒细胞的产生受到破坏。[24]癫痫病与疾病早期的新细胞产量增加,但异常整合有关,并在疾病晚期降低产量。[24]在癫痫发展过程中,成人生成的细胞异常整合可能会削弱齿状回阻止过度兴奋性活动到达海马锥体细胞的能力,从而促进癫痫发作。[24]持久性癫痫发作可刺激齿状颗粒细胞神经发生。这些新生的齿状颗粒细胞可能导致异常连接,从而导致与癫痫发生相关的海马网络可塑性。[25]
较短的颗粒细胞树突
患有阿尔茨海默氏症的患者的颗粒细胞树突较短。此外,与未患阿尔茨海默氏病的患者相比,树突的分支少,刺少。[26]然而,颗粒细胞树突不是老年斑的必需成分,并且这些斑对齿状回中的颗粒细胞没有直接影响。患有阿尔茨海默氏病,路易体变型和进行性核上性麻痹的患者出现齿状颗粒细胞的特定神经原纤维变化。[27]
参考
C. Reyher; J Liibke; W Larsen; G Hendrix; M Shipley; H Baumgarten (1991). "嗅球 Granule Cell Aggregates: Morphological Evidence for lnterperikaryal [sic] Electrotonic Coupling via Gap Junctions" (PDF). The Journal of Neuroscience. 11 (6): 1465–495. Retrieved 3 February 2016.
Llinas, Walton and Lang (2004). The Synaptic Organization of the Brain. Oxford University Press. doi:10.1093/acprof:oso/9780195159561.003.0007.
Claiborne BJ, Amaral DG, Cowan WM (1990). "A quantitative three-dimensional analysis of granule cell dendrites in the rat 齿状回". The Journal of Comparative Neurology. 302 (2): 206–219. doi:10.1002/cne.903020203. PMID 2289972.
David G. Amaral; Helen E. Scharfman; Pierre Lavenex (2007). "Progress in Brain Research - 齿状回: fundamental neuroanatomical organization". Progress in Brain Research. 163: 3–22. doi:10.1016/S0079-6123(07)63001-5. PMC 2492885. PMID 17765709.
Mugnaini E, Osen KK, Dahl AL, Friedrich VL Jr, Korte G (1980). "Fine 结构 of granule cells and related 中间神经元s (termed Golgi cells) in the 耳蜗 nuclear complex of cat, rat and mouse". Journal of Neurocytology. 9 (4): 537–70. doi:10.1007/BF01204841. PMID 7441303.
E. Young; O. Oertel (2004). The Synaptic Organization of the Brain. Oxford University Press. doi:10.1093/acprof:oso/9780195159561.003.0004.
K. Neville; L. Haberly (2004). The Synaptic Organization of the Brain. Oxford University Press. doi:10.1093/acprof:oso/9780195159561.003.0010.
V Egger; K Svoboda; Z Mainen (2005). "Dendrodendritic Synaptic Signals in 嗅球 Granule Cells: Local Spine Boost and Global Low-Threshold Spike". The Journal of Neuroscience. 25 (14): 3521–3530. doi:10.1523/JNEUROSCI.4746-04.2005. PMID 15814782.
Huang CM, Wang L, Huang RH (2006). "Cerebellar granule cell: ascending axon and 平行纤维". European Journal of Neuroscience. 23 (7): 1731–1737. doi:10.1111/j.1460-9568.2006.04690.x.
M Manto; C De Zeeuw (2012). "Diversity and Complexity of Roles of Granule Cells in the Cerebellar Cortex". The 小脑. 11 (1): 1–4. doi:10.1007/s12311-012-0365-7. PMID 22396329.
M. Bear; M. Paradiso (2006). Neuroscience: Exploring the Brain. Lippincott Williams & Wilkins. p. 855. ISBN 9780781760034.
P. Seja; M. Schonewille; G. Spitzmaul; A. Badura; I. Klein; Y. Rudhard; W. Wisden; C.A. Hübner; C.I. De Zeeuw; T.J. Jentsch (Mar 7, 2012). "Raising cytosolic Cl(-) in cerebellar granule cells affects their excitability and vestibulo-ocular learning". The EMBO Journal. 31 (5): 1217–30. doi:10.1038/emboj.2011.488. PMC 3297995. PMID 22252133.
Eccles JC, Ito M, Szentagothai J (1967). The 小脑 as a neural machine. Springer-Verlag. p. 56. doi:10.1007/978-3-662-13147-3.
Bengtssona, F; Jrntell, H (2009). "Sensory transmission in cerebellar granule cells relies on similarly coded mossy fiber inputs". PNAS. 106 (7): 2389–2394. doi:10.1073/pnas.0808428106. PMC 2650166. PMID 19164536.
A Arenz; E Bracey; T Margrie (2009). "Sensory re介绍s in cerebellar granule cells". Current Opinion in Neurobiology. 19 (4): 445–451. doi:10.1016/j.conb.2009.07.003. PMID 19651506.
Marr D (1969). "A theory of cerebellar cortex". The Journal of Physiology. 202 (2): 437–70. doi:10.1113/jphysiol.1969.sp008820. PMC 1351491. PMID 5784296.
M Colicos; P Dash (1996). "Apoptotic morphology of 齿状回 granule cells following experimental cortical impact injury in rats: possible role in spatial memory deficits". Brain Research. 739 (1–2): 120–131. doi:10.1016/S0006-8993(96)00824-4. PMID 8955932.
T Nakashiba; J Cushman; K Pelkey; S Renaudineau; D Buhl; T McHugh; V Rodriguez Barrera; R Chittajallu; K Iwamoto; C McBain; M Fanselow; S Tonegawa (2012). "Young Dentate Granule Cells Mediate Pattern Separation, whereas Old Granule Cells Facilitate Pattern Completion". Cell. 149 (1): 188–201. doi:10.1016/j.cell.2012.01.046. PMC 3319279. PMID 22365813.
Weedman DL, Ryugo DK (1996). "Projections from auditory cortex to the 耳蜗 nucleus in rats: synapses on granule cell dendrites". The Journal of Comparative Neurology. 371 (2): 311–324. doi:10.1002/(SICI)1096-9861(19960722)371:2<311::AID-CNE10>3.0.CO;2-V. PMID 8835735.
R Balu; R Pressler; B Strowbridge (2007). "Multiple Modes of Synaptic Excitation of 嗅球 Granule Cells". The Journal of Neuroscience. 27 (21): 5621–5632. doi:10.1523/JNEUROSCI.4630-06.2007.
Jansen, Jaclyn. "First glimpse of brain circuit that helps experience to shape perception". ScienceDaily. Retrieved 2 March 2014.
R Feil; J Hartmann; C Luo; W Wolfsgruber; K Schilling; S Feil; et al. (2003). "Impairment of LTD and cerebellar learning by 浦肯野细胞 specific ablation of cGMP-dependent protein kinase I." The Journal of Cell Biology. 163 (2): 295–302. doi:10.1083/jcb.200306148. PMC 2173527. PMID 14568994.
S Danzer; R Kotloski; C Walter; Maya Hughes; J McNamara (2008). "Altered morphology of hippocampal dentate granule cell presynaptic and postsynaptic terminals following conditional deletion of TrkB". 海马. 18 (7): 668–678. doi:10.1002/hipo.20426. PMC 3377475. PMID 18398849.
S Danzer (2012). "Depression, stress, epilepsy and adult neurogenesis". Experimental Neurology. 233 (1): 22–32. doi:10.1016/j.expneurol.2011.05.023. PMC 3199026. PMID 21684275.
J. Parent; T Yu; R Leibowitz; D Geschwind; R Sloviter; D Lowenstein (1997). "Dentate Granule Cell Neurogenesis Is Increased by Seizures and Contributes to Aberrant Network Reorganization in the Adult Rat 海马". The Journal of Neuroscience. 17 (10): 3727–3738. PMID 9133393.
Einstein G, Buranosky R, Crain BJ (1994). "Dendritic pathology of granule cells in Alzheimer's disease is unrelated to neuritic plaques" (PDF). The Journal of Neuroscience. 14 (8): 5077–5088. PMID 8046469.
Wakabayashi K, Hansen LA, Vincent I, Mallory M, Masliah E (1997). "Neurofibrillary tangles in the dentate granule cells of patients with Alzheimer's disease, Lewy body disease and progressive supranuclear palsy". Acta Neuropathologica. 93 (1): 7–12. doi:10.1007/s004010050576. PMID 9006651. |

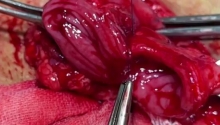
 因缺血而行小肠分段切除及一期吻合术
因缺血而行小肠分段切除及一期吻合术
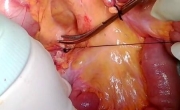
 肌肉活检 _ 如何进行肌肉活检
肌肉活检 _ 如何进行肌肉活检
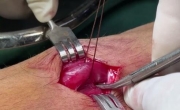
 尿道下裂修复后包皮瘘和龟头隐裂
尿道下裂修复后包皮瘘和龟头隐裂
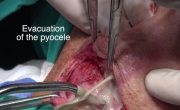 通过外部方法对喉囊肿进行有袋化或去顶
通过外部方法对喉囊肿进行有袋化或去顶

 发表于 2019-12-13 00:00:30
发表于 2019-12-13 00:00:30
 提升卡
提升卡 置顶卡
置顶卡 沉默卡
沉默卡 喧嚣卡
喧嚣卡 变色卡
变色卡 千斤顶
千斤顶 显身卡
显身卡