马上注册,结交更多好友,享用更多功能,让你轻松玩转社区。
您需要 登录 才可以下载或查看,没有账号?注册
×
锥体细胞或锥体神经元是在大脑区域(包括大脑皮层,海马和杏仁核)中发现的一种多极神经元。 金字塔形神经元是哺乳动物前额叶皮层和皮质脊髓束的主要兴奋单位。 金字塔形神经元也是死后狂犬病感染中发现特征性标志Negri体的两种细胞类型之一。[1] 金字塔形神经元最早是由圣地亚哥·拉蒙·卡哈尔[2] [3]发现和研究的。 从那时起,有关锥体神经元的研究就集中在从神经可塑性到认知的主题上。

通过高尔基体染色的人类新皮质锥体神经元。 请注意,顶端树突垂直于躯体上方延伸,而众多的基底树突则从细胞体的底部向侧面放射。

锥体细胞的重建。 体细胞和树突标记为红色,轴突乔木标记为蓝色。 (1)体(2)基底枝突(3)顶端树突(4)轴突(5)侧突轴突。
内容
1 结构
1.1 顶端树突
1.2 基底枝
1.3 树突棘
2 成长与发展
2.1 差异化
2.2 产后早期发展
3 信令
3.1 射击分类
4 功能
4.1 皮质脊髓束
4.2 认知
5 参考
结构

通过绿色荧光蛋白(gfp)可视化锥形神经元

海马锥体细胞
锥体神经元的主要结构特征之一是圆锥形的体细胞,即细胞体,其后被命名为神经元。锥体细胞的其他关键结构特征是单个轴突,一个大的顶端树突,多个基底树突以及存在树突棘。[4]
顶端树突
根尖树突从锥体细胞的躯体的顶点升起。顶端树突是一个单一的,长而厚的树突,随着与躯体的距离增加并向皮层表面延伸,其分支数次。[4]
基底枝
基底树突起于躯体的底部。基底树突树由三至五个初级树突组成。随着距躯体距离的增加,基底树突大量分支。[4]
锥体细胞是大脑中最大的神经元之一。无论是在人类还是啮齿动物中,锥体细胞体(体)的平均长度约为20μm。金字塔形树突的直径通常在半微米到几微米的范围内。单个枝晶的长度通常为数百微米。由于分支,锥体细胞的总树突长度可能达到几厘米。锥体细胞的轴突通常更长,并且广泛分支,总长度达到几厘米。
树突棘
树突棘接收进入锥体细胞的大多数兴奋性冲动(EPSP)。 1888年,Ramóny Cajal首先使用高尔基方法注意到了树突棘。拉蒙·卡哈尔(Ramóny Cajal)也是第一个提出增加神经元接受表面积的生理作用的人。锥体细胞的表面积越大,神经元处理和整合大量信息的能力就越大。躯体上没有树突棘,而树突棘的数量则逐渐增加。[3]在大鼠中,典型的顶端树突具有至少3,000个树突棘。人的顶端树突的平均长度约为大鼠的两倍,因此,人的顶端树突上存在的树突棘数量可能高达6,000。[5]
增长与发展
差异化
金字塔形发生在大脑早期发育期间。祖细胞在皮层下增殖性心室区(VZ)和脑室下区(SVZ)中被定位于神经元谱系。未成熟的锥体细胞经过迁移以占据皮质板,并在此进一步分化。内源性大麻素(eCBs)是一类分子,已被证明可指导脊髓细胞发育和轴突寻路[6]。转录因子如Ctip2和Sox5已被证明有助于锥体神经元引导轴突的方向。[7]
产后早期发展
已显示大鼠出生后早期的锥体细胞经历许多快速变化。在出生后的第3天和第21天之间,已显示出锥体细胞的大小增加了一倍,顶端树突的长度增加了5倍,而基础树突的长度增加了13倍。其他变化包括膜静息电位的降低,膜阻力的降低以及动作电位峰值的增加。[8]
发信号
像大多数其他神经元中的树突一样,树突通常是神经元的输入区域,而轴突是神经元的输出。轴突和树突都高度分支。大量的分支允许神经元向许多不同的神经元发送信号和从许多不同的神经元接收信号。
金字塔形神经元与其他神经元一样,具有许多电压门控离子通道。在锥体细胞中,树突中有大量的Na +,Ca2 +和K +通道,而在体细胞中则有一些通道。锥体细胞树突中的离子通道具有与锥体细胞体中相同离子通道类型不同的特性。亚阈值EPSPs和反向传播的动作电位激活了锥体细胞树突中的电压门控Ca2 +通道。锥体树突内动作电位的反向传播程度取决于K +通道。锥体细胞树突中的K +通道为控制动作电位的振幅提供了一种机制。[9]
锥体神经元整合信息的能力取决于它们接收的突触输入的数量和分布。单个锥体细胞接收约30,000个兴奋性输入和1700个抑制性(IPSP)输入。兴奋性(EPSPs)输入仅终止于树突棘,而抑制性(IPSPs)输入则终止于树突轴,躯体甚至轴突。金字塔形神经元可以被谷氨酸和GABA分别激活和抑制。[4]
射击分类
金字塔形神经元已根据其对400-1000毫秒电流脉冲的激发响应而分为不同的子类。这些分类是RSad,RSna和IB神经元。
RSad
RSad锥体神经元或适应正常的尖峰神经元会发射出单个动作电位(APs),其后是超极化后电位。持续时间后电位的增加会在神经元中产生尖峰频率适应(SFA)。[10]
RSna
RSna锥体神经元或非适应性规则尖峰神经元在脉冲后激发一系列动作电位。这些神经元没有显示出适应的迹象。[10]
IB
IB锥体神经元或内在爆发性神经元以2到5个快速动作电位的爆发响应阈值脉冲。 IB锥体神经元无适应。[10]
功能
皮质脊髓束
金字塔形神经元是皮质脊髓束中主要的神经细胞类型。正常的运动控制取决于皮质脊髓束中轴突与脊髓之间连接的发展。锥体细胞轴突遵循诸如生长因子之类的线索来建立特定的联系。通过正确的连接,锥体细胞会参与负责视觉引导的运动功能的电路。[11]
认识
前额叶皮层中的锥体神经元与认知能力有关。在哺乳动物中,锥体细胞的复杂性从后脑区到前脑区增加。锥体神经元的复杂程度可能与不同类人猿物种的认知能力有关。由于前额叶皮层从参与处理所有感觉方式的大脑区域接收输入,因此前额叶皮层内的锥体细胞似乎在处理不同类型的输入。[模糊]锥体细胞可能在皮层视觉处理区域内的复杂对象识别。[2]
另见
Cerebral cortex
Pyramidal tract
Chandelier cells - innervate initial segments of pyramidal axons
Rosehip neuron
参考
Sketchy Group, LLC. "2.3 rhabdovirus". SketchyMedical. Archived from the original on 2017-04-13.
Elston GN (November 2003). "Cortex, cognition and the cell: new insights into the pyramidal neuron and prefrontal function". Cereb. Cortex. 13 (11): 1124–38. doi:10.1093/cercor/bhg093. PMID 14576205.
García-López P, García-Marín V, Freire M (November 2006). "Three-dimensional reconstruction and quantitative study of a 锥体细胞 of a Cajal histological preparation". J. Neurosci. 26 (44): 11249–52. doi:10.1523/JNEUROSCI.3543-06.2006. PMID 17079652.
Megías M, Emri Z, Freund TF, Gulyás AI (2001). "Total number and distribution of inhibitory and excitatory synapses on hippocampal CA1 锥体细胞s". Neuroscience. 102 (3): 527–40. doi:10.1016/S0306-4522(00)00496-6. PMID 11226691.
Laberge D, Kasevich R (November 2007). "The apical dendrite theory of consciousness". Neural Netw. 20 (9): 1004–20. doi:10.1016/j.neunet.2007.09.006. PMID 17920812.
Mulder J, Aguado T, Keimpema E, et al. (June 2008). "Endocannabinoid signaling controls 锥体细胞 specification and long-range axon patterning". Proc. Natl. Acad. Sci. U.S.A. 105 (25): 8760–5. doi:10.1073/pnas.0803545105. PMC 2438381. PMID 18562289.
Fishell G, Hanashima C (February 2008). "Pyramidal neurons grow up and change their mind". Neuron. 57 (3): 333–8. doi:10.1016/j.neuron.2008.01.018. PMID 18255026.
Zhang ZW (March 2004). "Maturation of layer V pyramidal neurons in the rat prefrontal cortex: intrinsic properties and synaptic function". J. Neurophysiol. 91 (3): 1171–82. doi:10.1152/jn.00855.2003. PMID 14602839.
Magee J, Hoffman D, Colbert C, Johnston D (1998). "Electrical and calcium signaling in dendrites of hippocampal pyramidal neurons". Annu. Rev. Physiol. 60 (1): 327–46. doi:10.1146/annurev.physiol.60.1.327. PMID 9558467.
Franceschetti S, Sancini G, Panzica F, Radici C, Avanzini G (April 1998). "Postnatal differentiation of firing properties and morphological characteristics in layer V pyramidal neurons of the sensorimotor cortex". Neuroscience. 83 (4): 1013–24. doi:10.1016/S0306-4522(97)00463-6. PMID 9502243.
Salimi I, Friel KM, Martin JH (July 2008). "Pyramidal tract stimulation restores normal corticospinal tract connections and visuomotor skill after early postnatal motor cortex activity blockade". J. Neurosci. 28 (29): 7426–34. doi:10.1523/JNEUROSCI.1078-08.2008. PMC 2567132. PMID 18632946. |

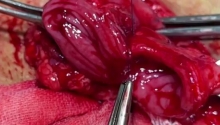
 因缺血而行小肠分段切除及一期吻合术
因缺血而行小肠分段切除及一期吻合术
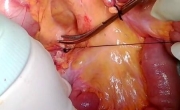
 肌肉活检 _ 如何进行肌肉活检
肌肉活检 _ 如何进行肌肉活检
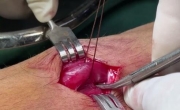
 尿道下裂修复后包皮瘘和龟头隐裂
尿道下裂修复后包皮瘘和龟头隐裂
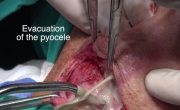 通过外部方法对喉囊肿进行有袋化或去顶
通过外部方法对喉囊肿进行有袋化或去顶

 发表于 2019-12-11 00:00:19
发表于 2019-12-11 00:00:19
 提升卡
提升卡 置顶卡
置顶卡 沉默卡
沉默卡 喧嚣卡
喧嚣卡 变色卡
变色卡 千斤顶
千斤顶 显身卡
显身卡