马上注册,结交更多好友,享用更多功能,让你轻松玩转社区。
您需要 登录 才可以下载或查看,没有账号?注册
×
本章的临床关键
妊娠和分娩期间发生的荷尔蒙和非荷尔蒙变化通过被称为胎盘单位的生理机制进行调节。 该单元的每个组件都产生一系列激素和发射器,它们在胎儿,胎盘和母体之间和之间具有多种效应。
胎儿胎盘单元的胎儿成分在妊娠和分娩的调节中起主要作用。 胎儿组分中的大多数活性发生在胎儿肾上腺中,其在妊娠中期比胎儿肾脏大。 胎儿肾上腺的胎儿区主要在胎儿期间分泌雄激素,这些雄激素作为胎盘中雌激素产生的前体。 胎儿肾上腺的总体作用尚未完全了解。
胎盘滋养细胞是人类促性腺激素(hCG)的来源,它可以在妊娠早期“抢救”黄体。 胎盘也产生大量的类固醇和肽类激素。 由于胎盘缺乏17α-羟化酶,它不能将黄体酮转化为雌激素。 胎盘替代使用来自胎儿肾上腺的雄激素作为孕前产生维持妊娠所需的雌激素。 除了雌激素,前列腺素和皮质类固醇,皮质醇,在胎盘中产生。 肽类激素包括人类生长激素(hPL),促肾上腺皮质激素释放激素(CRH)和催乳素。 其他重要的激素和传递物包括催产素,松弛素,前列腺素,白三烯和甲状旁腺激素相关肽。
母体对妊娠和分娩的适应和调节始于卵巢黄体分泌17-羟孕酮(17-OHP)。 如果母体卵巢缺乏黄体酮产生,怀孕可能会流产。 胎盘功能确保足够量的雌激素(主要是雌三醇或E3)增加子宫血流量以促进子宫生长,并且孕酮在妊娠期的大部分时间内维持子宫的静止状态。 肌层间隙连接的缺失促进了黄体酮的作用。
分娩过程(启动分娩过程)是从怀孕期间保持主动的功能性子宫静止状态中释放出来的。 这种静止在很大程度上是由于在分娩开始之前缺乏缝隙连接以及黄体酮的作用。 如下所述,分三阶段的其他三个阶段进入静止的第一阶段。 阶段1:通过子宫延伸和胎儿下丘脑 - 垂体 - 肾上腺(HPA)活动启动激活。 阶段2:刺激最可能始于胎盘产生CRH。 这个阶段继续宫颈成熟,子宫收缩力和蜕膜/胎儿膜激活。 阶段3:退化包括催产素释放急剧增加并且甲状旁腺激素相关肽(PTHrP)表达降低的胎儿排出。 该阶段还涉及胎盘分离和持续的子宫收缩。
妇女接受主要的内分泌和代谢改变,建立,维持和终止妊娠。 这些改变的目的是安全地输送能够在子宫外生存的婴儿。 胎儿的成熟和母亲的适应受到各种激素和传播者的调节(表5-1)。 本章涉及这些激素和非激素物质中最重要的属性,功能和相互作用,因为它们与怀孕和分娩有关。
胎儿胎盘单位
胎儿胎盘单位的概念是基于观察胎儿、胎盘和母体来源激素之间的相互作用。胎儿胎盘单位在很大程度上控制了妊娠的内分泌事件。虽然胎儿、胎盘和母亲都提供了输入,但胎儿似乎在其生长和成熟中发挥最活跃和控制的作用,并且可能在导致分娩的事件中发挥作用。

表 5-1
胎儿
肾上腺是胎儿的主要内分泌成分。在妊娠中期,胎儿肾大于胎儿。胎儿肾上腺皮质由外部决定性的或成人的区域和内部的、胎儿的区域组成。最终区域发育成成人肾上腺皮质的三个组成部分:束状带、肾小球带和网状带。在胎儿生活中,最终区主要分泌糖皮质激素和盐皮质激素。在胎儿期,胎儿区构成胎儿腺的80%,并主要分泌雄激素。它在出生后第一年就结束了分娩,完全消失了。胎儿肾上腺髓质合成并储存儿茶酚胺,在维持胎儿稳态中起重要作用。胎儿肾上腺在胎儿生长和成熟过程中的全部作用尚不完全清楚。
胎盘
胎盘在怀孕期间起着“额外的大脑”的作用,因为它包含了来自父亲和母亲的基因。此外,它是脑肽的来源,促肾上腺皮质激素释放激素(CRH),它在妊娠中具有非常重要的调节作用。因此,胎盘产生类固醇和肽类激素的量随孕龄的变化而变化。孕酮合成前体来源于母体循环。由于缺乏酶17α-羟化酶,人类胎盘不能直接将孕酮转化为雌激素,但必须使用雄激素,主要来自胎儿肾上腺,作为雌激素产生的前体来源。
母亲
母亲通过主要的内分泌和代谢变化适应妊娠。卵巢黄体产生孕酮(主要是17羟孕酮)在怀孕早期,直到其生产转移到胎盘。母体下丘脑和垂体后叶产生并释放催产素,引起子宫收缩和奶衰弱。垂体前叶产生催乳素,刺激乳汁分泌。母亲代谢的几个重要变化将在本章后面描述。
激素
胎儿胎盘单位产生各种激素以支持胎儿的成熟和母亲的适应。
肽激素
人绒毛膜促性腺激素
人绒毛膜促性腺激素(hCG)由胎盘的滋养层细胞分泌并维持妊娠。这种激素是一种分子量为40,000至45,000的糖蛋白,由两个亚基组成:α(α)和β(β)。 α亚基与黄体生成素(LH)和促甲状腺激素(TSH)共享。 hCG的特异性与其β亚基(β-hCG)相关,并且对β亚基具有特异性的放射免疫测定允许hCG的阳性鉴定。较新的hCG免疫测定法能够根据整个分子准确测量低水平的hCG,而不仅仅是β亚基。 hCG在妊娠以外的时间存在表明产生hCG的肿瘤,通常是葡萄胎,绒毛膜癌或胚胎癌(生殖细胞肿瘤)。
在怀孕期间,hCG在排卵后8天开始升高(中期LH峰值后9天)。这为几乎所有的免疫学或化学妊娠试验提供了基础。随着继续妊娠,hCG值在60至90天达到峰值,然后下降至中等,更恒定的水平。妊娠前6-8周,hCG维持黄体,从而确保孕酮持续输出,直到黄体酮生成转移到胎盘。 hCG滴度在异位妊娠或先兆子痫患者中可能异常低,而在滋养细胞疾病(如痣或绒毛膜癌)患者中异常高。这种激素还可以调节胎盘和胎儿肾上腺的类固醇生物合成,并刺激胎儿睾丸中的睾酮产生。早期妊娠的特点是调节性T细胞(Tregs)的增加,这种T细胞已知可以促进母体对胎儿的免疫耐受。最近的动物研究表明,hCG在怀孕期间充当这种免疫耐受的中枢调节剂。
人胎盘泌乳素
人胎盘泌乳素(hPL)起源于平面。它是一种分子量为22,300的单链多肽,在结构上类似于垂体生长激素和人催乳素。母体血清浓度平行胎盘重量,在整个妊娠期间上升到最后4周的最高水平。在足月,hPL占所有胎盘蛋白产量的10%。发现威胁性流产和胎儿宫内发育受限制的值较低。
人胎盘生乳素拮抗胰岛素的细胞作用并降低母体葡萄糖利用率,这增加了胎儿的葡萄糖利用率。这可能在糖尿病的发病机制中起作用。
促肾上腺皮质激素释放激素
在怀孕期间,CRH的主要来源是胎盘,它可以在妊娠12周时进入胎儿循环时进行测量。这种41个氨基酸的肽刺激胎儿肾上腺皮质激素(ACTH)的分泌,从而刺激胎儿肾上腺分泌硫酸脱氢表雄酮(DHEA-S),这是胎盘产生雌激素的重要前体物质。妊娠早期的胎儿肾上腺没有产生皮质醇的酶,但随着孕龄增加,它变得更有能力。胎儿皮质醇刺激胎盘CRH释放,然后刺激胎儿ACTH分泌,完成一个正反馈循环,在早产和足月的分娩和分娩过程中起着重要作用。已发现妊娠中期CRH水平升高与随后自发性早产的风险增加有关。
催乳素
催乳素是来自垂体前叶的肽,分子量约为20,000。正常的非怀孕水平约为10ng / mL。在妊娠期间,母亲催乳素水平上升,以响应增加母体雌激素的输出,刺激前垂体乳营养。催乳素的主要作用是刺激产后产奶。在妊娠的后半期,由胎儿垂体分泌的催乳素可能是胎儿肾上腺生长的重要刺激物。催乳素也可能在胎膜上的液体和电解质转移中起作用。
类固醇激素
黄体激素
黄体酮是最重要的人类孕激素。在黄体期,它会诱导子宫内膜的分泌变化,在怀孕期间,较高水平会诱导蜕膜变化。直到妊娠第六或第七周,黄体酮(17-羟孕酮)的主要来源是卵巢黄体。此后,胎盘开始发挥主要作用。如果在7周之前去除妊娠黄体并且需要继续妊娠,则应给予黄体酮以防止自然流产。循环的孕酮-1主要与载体蛋白结合,少于10%的游离和生理活性。
子宫肌层直接从排出胎盘的静脉血中接收黄体酮。 孕酮可预防子宫收缩,也可能参与建立受孕产品的免疫耐受。黄体酮还可抑制间隙连接形成,胎盘CRH表达以及雌激素,细胞因子和前列腺素的作用。因此,这种类固醇激素在整个怀孕期间维持子宫静止起着重要作用。
胎儿通过转化为皮质类固醇或通过羟基化或与惰性排泄产物结合而使孕酮失活。然而,胎盘可以将这些惰性材料转换回一号孕酮。类固醇生物化学途径如图5-1所示。
雌激素
胎儿和胎盘都参与雌酮,雌二醇和雌三醇的生物合成。胆固醇转化为胎盘中的孕烯醇酮。该前体主要在胎儿中转化为DHEA-S,在较小程度上转化为母体肾上腺。 DHEA-S进一步被胎盘代谢为雌酮(E1),并通过睾酮进一步代谢为雌二醇(E2)。雌三醇(E3)是人类妊娠中最丰富的雌激素,它是由胎儿肝脏中由DHEA-S产生的16α-羟基-DHEA-S在胎盘中合成的。在转化为E3之前,胎盘硫酸酯酶需要使16α-羟基-DHEA-S去偶联(图5-2)。胎盘中的类固醇硫酸酯酶活性很高,除了罕见的硫酸酯酶缺乏症。
雌激素在怀孕和分娩期间具有重要作用。它们增加子宫血流量,允许必要的子宫生长。它们有助于准备哺乳期的乳房组织,并刺激肝脏中激素结合球蛋白的产生。它们还与皮质醇一起在肺表面活性剂生产中起作用。
孕妇循环中雌三醇的突然下降可能表明胎儿在神经系统完整的胎儿中存在妥协。无脑胎儿缺乏下丘脑,垂体前叶和肾上腺发育不良;因此,雌三醇的产量只有正常的10%左右。
雄激素
在怀孕期间,雄激素主要来源于胎儿肾上腺皮质的胎儿区。 ACTH和hCG刺激雄激素分泌,后者主要在妊娠的前半期有效,当它存在于高浓度时。胎儿肾上腺倾向于通过睾酮和雄烯二酮产生DHEA。胎儿雄激素进入脐带和胎盘循环,并作为雌酮,雌二醇和雌三醇的前体(见图5-1)。
胎儿睾丸还分泌雄激素,特别是睾酮,其在靶细胞内转化为二氢睾酮(DHT),这是发育为男性外生殖器所必需的。 主营养刺激似乎是hCG。

图5-1类固醇激素生物合成的主要途径。 肾上腺DHEA主要作为其硫酸盐DHEA-S转运,DHEA-S也可以由硫酸胆固醇开始的类固醇硫酸盐形成。 LDL,低密度脂蛋白。

图5-2胎儿 - 胎盘单位中雌三醇的形成。 DHEA-S,脱氢表雄酮硫酸盐。
糖皮质激素
皮质醇源自循环胆固醇(见图5-1)。 母亲血浆皮质醇浓度在整个妊娠期间上升,并且是胎盘产生CRH的主要刺激因素。 在怀孕期间,皮质醇分泌的昼夜节律持续存在,除非患者具有显着的心理应激,这增加了晨间和傍晚的皮质醇水平,此时昼夜节律丧失。 妊娠期血浆转运蛋白水平升高,可能受雌激素刺激,无血浆皮质醇浓度增加一倍。
胎儿肾上腺和胎盘都参与皮质醇代谢。胎儿肾上腺由来自胎儿垂体的ACTH刺激产生皮质醇和DHEA-S。与胎儿区产生的DHEA-S相反,皮质醇来源于确定区(见图5-1)。在妊娠结束时,皮质醇促进II型肺泡细胞的分化和表面活性剂的生物合成和释放进入肺泡。表面活性剂降低了肺部膨胀所需的力量。表面活性剂的不足导致早产儿的呼吸窘迫,这可能导致死亡。皮质醇还在激活分娩中起重要作用,增加胎盘CRH和前列腺素的释放。
其他激素和发射器
催产素
产生于母体下丘脑的视上核和室旁核的催产激素原沿着神经纤维向下迁移,催产素在垂体后叶的神经末梢积聚。催产素是通过各种刺激从垂体后叶释放的九肽,如产道扩张和乳腺刺激。催产素导致子宫收缩,但催产素产生受损,如在尿崩症中,不影响正常分娩。分娩前催产素循环水平的波动与子宫活动的变化不一致。母亲血清催产素水平仅在第一产程中上升。催产素可用于诱导分娩,特别是在中期妊娠中,或增加自发分娩过程中的收缩频率和强度。
最近,催产素已经显示出影响涉及情绪,认知和社交行为的大脑区域。对“亲社会”行为的影响包括对放松,信任和心理稳定性的积极影响。如果得到证实,这些影响可能有助于分娩,分娩和善后护理。
松弛素
松弛素是一种起源于卵巢黄体的肽激素。胎盘也会产生松弛素,并在妊娠第10周达到母体循环中的最高浓度,然后下降。松弛素与子宫颈的软化有关,这是妊娠的一种明显的体征。其主要功能似乎是通过促进血管生成来促进胚胎的植入。在进行体外受精(IVF)的妇女的卵巢过度刺激期间,卵巢产生过量的松弛素。已显示松弛素过量与宫颈缩短和早产风险增加有关。
前列腺素和白三烯
前列腺素是一种普遍存在的,具有生物活性的脂质,涉及广泛的生理和病理生理反应。它们不是真正的激素,因为它们不是在一个腺体中合成并通过循环血液运输到目标器官。相反,它们在其作用地点或附近合成。前列腺素E2(PGE2)和前列腺素F2α(PGF2α),前列环素和血栓素A2在子宫内膜,子宫肌层,胎膜,蜕膜和胎盘中合成。 PGE2和PGF2α引起子宫收缩。它们在子宫肌层中的受体在怀孕期间下调。前列腺素还可以引起其他平滑肌的征象,例如肠道的平滑肌。因此,当在药理学上使用时,前列腺素可能引起不希望的副作用,例如恶心,呕吐和腹泻。 PGE2和PGF2α的羊水浓度在整个怀孕期间上升并在自发分娩过程中进一步增加。需要催产素引产的女性比接受自愿分娩的女性要低。通过各种途径施用PGE 2或PGF2α在妊娠的任何阶段诱导分娩或流产。目前正在使用各种合成前列腺素衍生物来终止任何阶段的妊娠并在足月时间诱导分娩。
前列腺素被认为在启动和控制劳动力方面发挥重要作用。 前列腺素合成始于花生四烯酸的形成,花生四烯酸是“2”系列前列腺素的必需前体(即PGE2,PGF2α)。 花生四烯酸以酯化形式作为甘油磷脂储存在滋养层细胞膜中。 起始步骤是磷脂酶A2或C催化的甘油磷脂水解。磷脂酶A2优先作用于绒毛膜磷脂酰乙醇胺释放花生四烯酸(图5-3)。 游离的花生四烯酸的酸不会积累。 劳动似乎伴随着绒毛膜,羊膜和蜕膜中的一系列事件,它们从储存的形式释放花生四烯酸并将其转化为活性前列腺素。 17β-雌二醇刺激几种从花生四烯酸的酸合成前列腺素中有活性的酶。

图5-3前列腺素和白三烯生物合成图。
有两种称为COX-1或PGHS-1的环加氧酶同工酶,以及COX-2或PGHS-2。 这些同工酶来自不同的基因。 COX-1在静息细胞中表达,而COX-2是可诱导的。 它在细胞活化时在炎症部位表达,并加强炎症过程。
胎膜中COX-1 mRNA表达低
而且不随胎龄而改变,而羊膜中COX 2 mRNA的表达随着孕龄而增加。
磷脂酶A2活性增加可能导致早产。 宫颈内,宫内或尿道感染往往与早熟劳动有关。 许多产生这些感染的生物体具有磷脂酶A2活性,其可以产生游离的花生四烯酸,然后是前列腺素合成,这可能引发分娩。
前列腺素合成酶抑制剂可延长妊娠期。非甾体类抗炎药(NSAIDs)抑制磷脂酶A2,而阿司匹林类药抑制环氧合酶。因为PGE2使动脉导管保持开放,所以在摄入大量NSAID或阿司匹林或长时间摄入导管后可能会过早闭合导管,导致胎儿肺动脉高压和死亡。
花生四烯酸代谢的另一途径是将花生四烯酸转化为白三烯(见图5-3)。前列腺素和白三烯均诱导蜕膜化,这意味着它们在妊娠早期引发子宫内膜的变化以促进受精卵的植入。
尽管PGF2α在产生子宫收缩活性方面更有效,但PGE2是通过诱导结缔组织变化使子宫颈成熟的最有效的前列腺素。因此,PGE2及其合成衍生物在诱导分娩或流产之前临床上用于宫颈成熟。
母体代谢的变化
如下所述,母体代谢通过内分泌调节适应妊娠。
血管紧张素 - 醛固酮
醛固酮是在肾上腺皮质的肾小球区中合成的盐皮质激素。怀孕的主要来源是母亲肾上腺。虽然胎儿肾上腺能够合成它,但胎儿肾上腺和胎盘并没有显著参与醛固酮的产生。醛固酮分泌受肾素 - 血管紧张素系统调节。肾脏中形成的肾素增加将血管紧张素原(肾素 - 底物)转化为血管紧张素I,血管紧张素I进一步代谢为血管紧张素II,血管紧张素II又刺激醛固酮分泌。醛固酮刺激肾脏的远端小管中钠的吸收和钾的分泌,从而维持钠和钾的平衡。妊娠期间,肾素 - 底物(一种血浆蛋白)的浓度升高。据认为,怀孕期间存在的孕激素和雌激素的高浓度刺激肾素和肾素 - 底物形成,从而导致血管张力素II水平增加和醛固酮产生增加。妊娠高血压的醛固酮分泌率下降,在某些情况下,可能会降至非妊娠水平。
钙代谢
虽然妊娠期的钙吸收增加,但总母体血清钙下降。总钙的下降与血清白蛋白的下降相似,因为大约一半的钙与白蛋白结合。由于母体甲状旁腺激素的产生增加,离子钙(生理上重要的钙部分)在整个妊娠期间基本上保持稳定。后者促进钙通过胎盘转移到胎儿以获得充足的骨骼发育,同时从母体骨骼中调动钙以维持足够的钙稳态。在妊娠后期,与胎儿骨骼的最大钙化相一致,血清甲状旁腺激素水平升高增加了母亲钙吸收和骨吸收。
钙离子通过胎盘主动转运,妊娠晚期胎儿血清总血清水平和离子钙水平均高于母体水平。高胎儿离子钙抑制胎儿甲状旁腺激素的产生,甲状旁腺激素不会穿过胎盘。此外,刺激降钙素的产生,从而为胎儿提供足够的钙,用于钙化钙的形成。在产后的前24至48小时,新生儿的总血清钙浓度通常下降,而磷浓度上升。两者都在1周内适应成人水平。
分娩
分娩意味着分娩,劳动是胎儿从子宫排出到外界的生理逻辑过程。
生物化学收缩的基础
肌动蛋白收缩是由腺苷三磷酸(ATP)和钙驱动的肌动蛋白和肌球蛋白丝的滑动引起的。尽管骨骼肌需要神经支配,但平滑肌如子宫肌层的收缩主要由激素刺激引发。在子宫肌细胞膜中发现了激素受体。

图5-4肌醇三磷酸酯形成图。
催产素和前列腺素与它们各自的受体的结合激活磷脂酶C,磷脂酶C水解存在于细胞膜中的脂质磷脂酰肌醇二磷酸,形成肌醇三磷酸酯和二酰甘油(图5-4)。肌醇三磷酸酯诱导钙从肌质网释放,细胞内钙储存区域。由此产生的高细胞内游离钙浓度使得子宫肌层的肌原纤维收缩。随后,钙在ATP的帮助下被泵回到肌浆网状结构中,并且更多的钙可以通过短暂打开的电压操作和受体操作通道从细胞外液进入。维持足够的母体钙水平很重要,因为在剖宫产分娩风险的妇女中观察到孕妇血清钙水平低。
与其中存在His束的心脏不同,在子宫中未发现收缩同步的解剖结构;然而,最近观察到怀孕期间维生素D缺乏与肌功能障碍和剖宫产分娩风险更高有关。子宫收缩随着电流从细胞流向低阻力区域而扩散。这些区域与间隙连接相关,这在分娩时变得尤为突出。雌二醇和前列腺素促进间隙结合的出现,而黄体酮反对雌二醇的这种作用。
妊娠期激素控制与分娩发动
妊娠期长度受胎儿的荷尔蒙控制。每个物种不仅具有独特的妊娠长度,而且还具有控制该长度的独特机制。因此,虽然动物模型提供了重要的见解,但它们没有提供关于控制人体形态长度的具体信息或控制开始劳动的机制。
动物模型
大多数研究都是在绵羊身上进行的,胎儿似乎控制着分娩的开始。胎儿下丘脑刺激胎儿垂体分泌促肾上腺皮质激素,从而导致胎儿肾上腺皮质醇的激增。皮质醇激增诱导胎盘酶17α-羟化酶和雌激素前体的雄激素形成(参见图5-1),同时降低孕酮形成。雌激素与孕激素比例的增加导致(1)前列腺素分泌增加; (2)形成子宫肌层间隙结构,提供对电流低阻力区域并增加协调子宫收缩的区域; (3)宫颈成熟;和(4)劳动的开始。给予ACTH,糖皮质激素或地塞米松也可以引发分娩。去除胎儿垂体或肾上腺,这两者都是皮质醇激增所必需的,将导致妊娠过长。
在具有导致胎儿垂体和肾上腺功能障碍的遗传缺陷的一群格恩西奶牛中,怀孕延长,并且不发生正常的阴道分娩。在兔子中,分娩直接导致黄体酮产生的下降,其次是黄体功能下降。通过服用黄体酮可以预防流产。
人类
基于动物和人类的研究,正常的自发人类分娩过程可分为四个阶段。
阶段0:静止。在整个怀孕期间,子宫保持相对静止状态。在怀孕期间,肌肉活动受到各种物质的抑制,但孕酮似乎在维持子宫静止中发挥中心作用。最近,已经表明,各种器官如肺,心脏,膀胱和子宫受到在每个器官中产生的甲状旁腺激素相关肽(PTHrP)的调节。这种肽类激素受到拉伸刺激,并且在妊娠期间,PTHrP可使子宫松弛以促进胎儿生长。分娩时,调节PTHrP的基因被关闭,允许子宫收缩并开始退化过程。这减少了产后出血的风险。在静止期发生的罕见子宫收缩频率和幅度较低,协调性较差;这些通常被称为女性Braxton-Hicks收缩。这些收缩不协调主要是由于怀孕的子宫肌层缺乏缝隙连接。
阶段1:激活。正常情况下,肌层激活的信号可能来自胎儿生长导致的子宫延伸,或胎儿成熟后胎儿下丘脑 - 垂体 - 肾上腺(HPA)轴的激活。在动物模型中已显示子宫伸展以增加子宫肌层中的间隙连接和收缩相关蛋白。目前认为,一旦达到胎儿成熟度(由尚未知的机制决定),胎儿下丘脑可增加CRH分泌,这反过来又刺激胎儿垂体和皮质醇引起的ACTH表达和胎儿的雄激素产生肾上腺。最近来自怀孕小鼠的数据表明,胎儿通过将主要肺表面活性蛋白SP-A分泌到羊水中来表明胎儿开始分娩。
这些数据支持胎儿HPA轴在分娩开始时的关键作用,因为表皮蛋白合成受到糖皮质激素的刺激。 PTHrP的作用也可能在肺部发育和分娩发作中起重要作用。 胎儿肺在分娩初期的作用的概念特别有吸引力,因为胎儿肺是最后一个主要器官成熟。
阶段2:刺激。阶段2涉及导致分娩的共同途径的逐步级联事件,涉及子宫收缩,宫颈成熟和蜕膜/胎膜激活。
这种级联可能始于胎盘产生CRH。与糖皮质激素对母体下丘脑CRH合成的抑制作用相反,胎盘CRH合成受到糖皮质激素的刺激。胎盘CRH进入胎儿循环,反过来促进胎儿皮质醇和DHEA-S的产生。这种正反馈循环逐渐被放大,从而推动从胎儿HPA激活到分娩和胎盘生产雌激素的过程。
对于大多数的怀孕,子宫静止主要由孕激素的作用。在大多数哺乳动物怀孕结束时,母体黄体酮水平下降,雌激素水平上升。在人类和非人类灵长类妊娠中,孕酮和雌激素的浓度在整个怀孕期间继续增加,直到胎盘分娩。通过改变孕激素受体(PR)的表达,女性和非人灵长类动物可能出现功能性黄体酮戒断症状。人类子宫肌层中有两个孕酮受体(PRA和PRB)。与增加黄体酮作用的PRB相反,PRA抑制黄体酮作用。分娩时子宫肌层中PRA与PRB的比例增加,这实际上导致了促性腺激素撤回。
功能性黄体酮戒断导致功能性雌激素占优势,部分原因是雌激素胎盘产生增加。雌激素受体(ER)异构体ERα的表达通常被孕酮抑制,但随着PR-A的表达相对于PR-B的表达增加,在产褥子肌组织中ERα的表达也增加。 ERα的表达上升有助于增加雌激素作用。增加雌激素的水平也增强许多雌激素依赖性收缩相关蛋白(CAP)的表达,包括连接蛋白43(间隙连接),催产素受体,前列腺素受体,环氧合酶-2(COX-2,导致前列腺素产生)和肌球蛋白轻链激酶(MLCK,刺激肌层收缩性和分娩)。
生物过程的逐步级联导致分娩的共同途径,涉及宫颈成熟,子宫收缩力和蜕膜/胎膜激活。宫颈成熟主要由前列腺素的作用,间隙连接和肌球蛋白轻链激酶作用下的子宫收缩性,以及金属蛋白酶等酶的作用导致的蜕膜/胎膜膜激活,最终导致细胞破裂膜。
第3阶段:参与。在驱逐胎儿期间,产后催产素的释放急剧增加,这有助于开始最后阶段的分娩。据认为,催产素的一些增加来自胎儿脑垂体,这是胎儿头部通过骨盆时受压的结果。在胎儿从子宫排出后的短时间内,PTHrP的作用下降,这促进了正常的非妊娠子宫收缩恢复。这种减少说明子宫对催产素的敏感性增加。阶段3涉及胎盘分离和继续子宫收缩。通过沿着蜕膜基底面的切割发生胎盘分离。子宫收缩对预防胎盘分娩后暴露的大静脉窦出血至关重要,主要受催产素影响。在早期母乳喂养期间,这通过催产素的降低得到了进一步的支持。
参考:hacker & moore’s essentials of obstetrics & gynecology---6e |

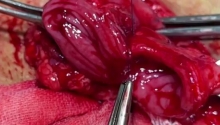
 因缺血而行小肠分段切除及一期吻合术
因缺血而行小肠分段切除及一期吻合术
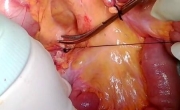
 肌肉活检 _ 如何进行肌肉活检
肌肉活检 _ 如何进行肌肉活检
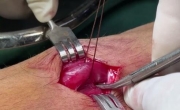
 尿道下裂修复后包皮瘘和龟头隐裂
尿道下裂修复后包皮瘘和龟头隐裂
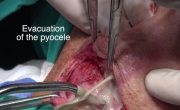 通过外部方法对喉囊肿进行有袋化或去顶
通过外部方法对喉囊肿进行有袋化或去顶

 发表于 2018-6-18 12:47:30
发表于 2018-6-18 12:47:30
 提升卡
提升卡 置顶卡
置顶卡 沉默卡
沉默卡 喧嚣卡
喧嚣卡 变色卡
变色卡 千斤顶
千斤顶 显身卡
显身卡