马上注册,结交更多好友,享用更多功能,让你轻松玩转社区。
您需要 登录 才可以下载或查看,没有账号?注册
×
本章的临床关键
女性生殖周期(月经周期)可以被看作是四个独立的生理周期(下丘脑,垂体,卵巢和子宫内膜),但实际上是一个非常复杂和整合的事件。 这28(±7)天的周期允许卵子成熟和释放(通常只有一个),如果受精可以植入接受性子宫内膜。 如果没有发生受精和植入,最终的结果就是月经。
下丘脑,它释放促性腺激素释放激素(GnRH)的脉冲和垂体腺之间的沟通是必要的,以允许垂体对卵巢激素(雌二醇和孕酮)和其他因素的变化作出反应。 这种重要的沟通结果对于垂体性腺激素,黄体生成激素(LH)和卵泡刺激素(FSH)的释放产生负面和正面的反馈。 这些促性腺激素也以脉冲形式释放,刺激卵巢中的卵泡生长(FSH),使得一个优势卵泡释放卵母细胞以响应LH的中周期激增。 其他生长因子和肽如抑制素-A,抑制素-B和激活素在全身和局部起作用以控制卵泡生长。
卵母细胞释放后,显性(graafian)卵泡成为黄体并分泌雌二醇(E2)和黄体酮(P4)。 在排卵前,子宫内膜对E2具有生长或增殖的反应,并且在排卵后具有成熟的允许植入的P4和E2,如果受精发生的话。 如果发生妊娠,黄体继续保持子宫内膜,由胎盘组织中的人绒毛膜促性腺激素(hCG)刺激。
受精和植入开始于正常的精子功能和输卵管内的卵母细胞的渗透。受精恢复染色体的二倍体数目并决定受精卵的性别。受精卵在排卵后3天内到达子宫内膜,并在数天之后到达胚泡植入物。
当胚泡进入子宫内膜时,滋养层束分支形成原始绒毛。绒毛在受精后的第十二天首先表现出来,是胎盘的基本结构。胎盘是妊娠期胎儿发育的生命支持系统。
生殖周期
每个女性生殖周期(月经周期)代表了低丘脑,垂体,卵巢和子宫内膜之间的复杂相互作用。 促性腺激素,黄体生成素(LH)和促卵泡激素(FSH)以及性类固醇激素(主要是雌二醇(E2)和孕酮(P4))的周期性变化诱导了卵巢的功能和形态学变化,导致卵泡成熟,排卵和黄体形成。 在子宫内膜水平上的类似变化允许成功植入受精卵或在未发生早孕时生理性地排出月经子宫内膜。 按照惯例,正常周期从月经出血的第一天开始,并在下一次月经的第一天之前结束。 每个周期的平均长度为28(±7)天。
生殖周期可以从四种生理成分的角度来看待(表4-1)。 在本章中,依次接近下丘脑 - 垂体轴,卵巢和子宫内膜的周期性变化,好像它们是四个不同的周期一样,但这些内分泌事件以独特的整合方式发生。 本章还包括关于精子发生,受精,植入和胎盘的讨论。

表 4-1 
生殖(月经)周期的内分泌组分
下丘脑 - 垂体轴
垂体
脑下垂体位于骨底部的下丘脑下方(蝶鞍),并通过鞍鞍上的硬脑膜(鞍膈)的冷凝而从颅腔分离。垂体分为神经垂体和腺垂体两个主要部分(图4-1)。神经垂体由后叶(神经部分)、神经柄(漏斗部)和正中隆起组成,起源于神经组织,与下丘脑和中枢神经系统有直接的连续性。腺垂体包括外胚叶(前叶)、中间间叶(中间叶)和围绕神经柄的结节部,是由外胚层衍生而来的。
中间隆起和神经柄(垂体门系统)的动脉血供应是下丘脑分泌到垂体前叶的主要途径。
神经垂体主要作用于从下丘脑视上和脑室旁核的神经元投射向其循环释放催产素和抗利尿激素(抗利尿激素)。
垂体前叶含有不同的细胞类型,产生六种蛋白激素:促卵泡激素(FSH)、促黄体生成激素(LH)、促甲状腺激素(TSH)、催乳素、生长激素(GH)和促肾上腺皮质激素(ACTH)。
促性腺激素,FSH和LH,合成和存储在细胞中称为促性腺激素,而TSH是由促甲状腺激素产生的。FSH、LH和TSH是由α和β亚基组成的糖蛋白。FSH、LH和TSH的α亚基是相同的。人绒毛膜促性腺激素(HCG)中也存在相同的α亚基。β亚基是每个激素的个体。循环LH的半衰期约为30分钟,而FSH的半衰期为数小时。半衰期的差异可能至少部分地说明了这两种促性腺激素的差异分泌模式。

图4-1下丘脑-垂体-门脉循环系统。弓状核中促性腺激素释放激素(GnRH)的脉冲通过这种循环系统输送到垂体前叶。GnRH脉冲的中断或显著改变将导致生殖周期停止。
催乳素是由乳糖分泌的。与腺垂体分泌的其他肽激素不同,催乳素的垂体释放受到下丘脑的紧张性抑制。催乳素循环的半衰期约为20至30分钟。除了催乳素的作用外,催乳素可能直接或间接地影响下丘脑、垂体和卵巢功能,与排卵周期有关,尤其是在慢性高尿酸-泌乳素血症的病理状态下(见第33章)。
促性腺激素分泌细胞模型
正常卵巢周期可分为卵泡期和黄体期(图4-2)。卵泡期开始于月经的开始,并在LH排卵前期达到高潮。黄体期开始于排卵前LH波峰的开始,并在月经的第一天结束。
从前一个周期的黄体退化降低雌二醇和孕酮通过负反馈机制引起FSH升高,刺激卵泡生长和雌二醇分泌。卵泡生长和雌二醇分泌的一个主要特征是由两个促性腺激素(LH和FSH)两个细胞(CeA和Grunu LuSa细胞)的卵巢卵泡发育理论解释。根据这一理论,在卵泡中有单独的细胞功能,其中LH刺激卵泡膜细胞产生雄激素(雄烯二酮和睾酮),然后促卵泡素细胞将这些雄激素转化为雌激素(雄烯二酮,雌酮和T)。如图4-3所示,雌二醇与雌二醇。最初,在较低水平的雌二醇中,从垂体促性腺激素中的促性腺激素池释放LH有负反馈作用。当卵泡期后期雌二醇水平升高(>200皮克>50小时)时,促性腺激素释放呈正相关,导致LH激增和排卵。后者发生36至44小时后,这中间周期LH浪涌。随着避孕药中含有的药物剂量的增加,对促性腺激素释放激素(GnRH)有着深刻的负反馈作用,从而没有释放促性腺激素池。因此排卵被阻断(见第27章)。
在黄体期,LH和FSH均通过升高的雌二醇和孕酮的负反馈作用而被显著抑制。这种抑制一直持续到黄体期结束时黄体期黄体期的孕酮和雌二醇水平下降,如果妊娠不能发生。净效应是血清FSH的轻微上升,这为下一个周期启动了新的卵泡生长。黄体功能回归的持续时间是这样的,月经一般发生在LH激增后14天,在没有怀孕的情况下。当妊娠发生时,黄体被HCG“拯救”,其作用如LH,孕激素继续生产。
日常周期

图4-2正常月经周期中的激素活性和水平。 图中显示了周期的所有四个组成部分(下丘脑,垂体,卵巢和子宫内膜)。 这些组件以高度整合的方式工作,代表了生理学中最复杂的内分泌/终末器官系统之一。 FSH,促卵泡激素; GnRH,促性腺激素释放激素; LH,黄体生成素。
下丘脑
从下丘脑分离出五种影响生殖周期的小肽或生物胺。所有这些对垂体前叶的激素分泌都有特定的影响。促性腺激素释放激素(GnRH)、促甲状腺素释放激素(TRH)、生长激素释放抑制因子(SRIF)或生长抑素、促肾上腺皮质激素释放因子(CRF)和催乳素释放抑制因子(PIF)。本章仅讨论GnRH和PIF。
GnRH是一种主要在弓形核中合成的十肽。它的半衰期很短,只有2到4分钟。它负责LH和FSH的合成和释放。因为它通常导致释放更多的LH比FSH,它通常不被称为LH释放激素(LH-RH)或LH释放因子(LRF)。FSH和LH在垂体-性腺-滋养体中以两种不同的形式存在。一种是可释放形式,另一种是存储形式。GnRH通过垂体门血管到达垂体前叶并刺激FSH和LH的SYN论文,它们被储存在促性腺激素中。随后,GnRH激活并将这些分子转化为可释放的形式。促性腺激素释放激素也可诱导LH和FSH立即释放到循环中。GnRH受体已在卵巢等其他组织中发现,GnRH也可能对卵巢功能有直接的影响。
GnRH在整个月经周期中以脉冲方式分泌,如图4-4所示。通过测量LH脉冲间接评估GnRH释放的频率在个体间差异很大。在卵泡期GnRH脉冲更频繁,振幅更低,而在黄体期,脉冲发生的频率较低,但振幅更高。

图4-3两个促性腺激素(LH和FSH),两个细胞(卵泡膜顶细胞和颗粒细胞)卵泡发育的理论。每个细胞被理论化为单独的功能;LH刺激在卵泡膜细胞中产生雄激素(雄烯二酮和睾酮),FSH刺激这些雄激素在颗粒细胞中对雌激素(雌酮和雌二醇)的芳构化。促卵泡激素、促卵泡素、促黄体生成素、促黄体生成素。
静脉注射和皮下注射外源性脉冲GnRH已被用于诱导排卵在选定的妇女谁不排卵作为下丘脑功能障碍的结果。GnRH的连续(非搏动)灌注可通过垂体促性腺激素的“下调”或脱敏过程可逆地抑制促性腺激素分泌。这代表了GnRH类似物(包含九个氨基酸的非肽类)的基本作用机制,它成功地用于治疗卵巢激素依赖性疾病如子宫内膜异位症、平滑肌瘤(纤维瘤)、多毛症和性早熟。促性腺激素释放激素(GnRH)既有激动作用又有拮抗作用。
几种机制控制GnRH的分泌。
雌二醇似乎增强GnRH的下丘脑释放,并且可能通过增加GnRH释放或通过增强对十肽的脑垂体响应而帮助诱导中周期LH刺激。 促性腺激素对GnRH释放具有抑制作用。 儿茶酚胺也可能起着主要的调节作用。 多巴胺在弓形和脑室周核中合成,并且可能通过项目的结节漏斗部的道对GnRH分泌具有直接抑制作用到正中隆起。 5-羟色胺也似乎抑制GnRH脉冲释放,而去甲肾上腺素刺激它。 内源性阿片类药物可以部分受卵巢类固醇调节的方式抑制GnRH从下丘脑的释放。
促性腺激素释放激素脉冲

图4-4促性腺激素释放激素(GnRH)在正常月经周期中的脉冲释放。
下丘脑产生PIF,其对乳糖释放的催乳素释放具有慢性抑制作用。许多影响多巴胺能机制的药理学药物(例如氯丙嗪)也影响催乳素释放。多巴胺本身被下丘脑神经元分泌到垂体门静脉血管中,并直接抑制垂体内催乳素的释放。基于这些观察,已经提出下丘脑多巴矿可能是主要的PIF。除了由PIF调节催乳素释放外,下丘脑也可能产生催乳素释放因子(PRF),在某些条件下,催乳素释放可引起大而迅速的增加,例如哺乳期的乳房刺激。PIF和PRF都没有生物化学特征明确为2014。TRH也起到催乳素释放的作用。这种表型不可解释原发性甲状腺功能减退症(继发性TRH升高)与高催乳素血症之间的关联。GnRH的前体蛋白,称为GnRH相关肽(GAP),已被鉴定为催乳素分泌的有力抑制剂和促性腺激素释放的增强子。这些结果表明,这种GnRH相关肽也可以是生理性PIF,并且可以解释在许多生殖状态中促性腺激素和催乳素分泌之间的反比关系。
卵巢周期
雌激素类
在卵泡发育早期,循环雌二醇水平相对较低。排卵前大约1周,水平开始增加,开始缓慢,然后迅速。在卵泡颗粒细胞中睾酮向雌二醇的转化是通过一种被称为芳构化的酶过程发生的,如图4-3所示。水平通常在中环LH峰之前达到最大1天。在这个高峰和排卵前,有一个明显而陡峭的瀑布。在黄体期,雌二醇在排卵后5至7天达到最大值,并在月经前不久回到基线。卵巢分泌的雌酮(E1,较弱的雌激素)明显低于雌二醇的分泌,但遵循相似的模式。雌酮主要来源于雄烯二酮通过芳香化酶的作用转化(图4-5)。
孕激素类
卵泡发育过程中,卵巢仅分泌少量孕酮和17α-羟孕酮。大部分的孕激素来自肾上腺孕烯醇酮和孕烯醇酮硫酸盐的外周转化。在排卵前,未破裂但黄体化的卵泡开始产生越来越多的孕激素睾酮。此时,血清17α-羟孕酮也显著增加。基础体温的升高在时间上与孕酮的中心效应有关。与雌二醇一样,黄体的孕激素分泌在排卵后5至7天达到最大值,并在月经前不久恢复到基线。如果怀孕发生,孕酮水平,因此基础体温,保持升高。
雄激素类
卵巢和肾上腺都分泌少量睾酮,但睾酮大部分来自雄甾酮的代谢,雄甾酮也由卵巢和肾上腺分泌。接近中环,血浆雄雄二酮的增加,这反映了增强的毛囊分泌。在黄体期,第二次上升发生在雄烯二酮,这反映了增强的黄体分泌。肾上腺也分泌雄甾酮,其昼夜模式与皮质醇相似。卵巢分泌少量的非常有效的二氢睾酮(DHT),但大部分的DHT源自雄烯二酮和睾酮的融合。大多数脱氢表雄酮(DHEA)和几乎所有的DHEA硫酸酯(DHEA S),是弱雄激素,由肾上腺分泌,尽管少量的DHEA由卵巢分泌。

图4-5甾体途径显示红色中的芳构化。Cmpd B,皮质酮;CMPD S,II脱氧皮质醇;脱氢表雄酮,脱氢表雄酮;DOC,脱氧皮质酮;OH,羟化酶。
血清结合蛋白
循环雌激素和雄激素大多与特定的性激素结合球蛋白(SHBG)或血清白蛋白结合。性激素的其余部分是游离的(游离的),这是生物逻辑活性部分。目前尚不清楚是否与血清蛋白(例如白蛋白)结合的类固醇可用于组织摄取和利用。SHBG在肝脏中的合成增加了雌激素和甲状腺激素,但睾酮降低。
催乳素
血清催乳素水平不明显变化在正常月经周期。催乳素和催乳素释放的响应于TRH的血清水平在黄体期比在周期的卵泡期稍高。这表明,大量的循环雌二醇和孕酮可增强催乳素释放。催乳素释放在白天变化,在睡眠过程中发生的最高水平。
催乳素可能参与卵巢类固醇生成的控制。卵泡液中催乳素浓度在卵泡发育过程中明显变化。催乳素浓度最高的小卵泡在早期卵泡期。卵泡液中催乳素浓度可能与孕酮的生成呈负相关。此外,高催乳素血症可能改变促性腺激素分泌。尽管这些观察,在正常月经周期中的促乳素的生理作用尚未明确建立。
卵泡发育
妊娠20周时胎儿卵母细胞数目最多,为6~700万。卵母细胞的显著闭锁(生理丧失)发生在出生时,在卵巢中只剩下1到200万个。青春期(闭锁期)300000~400000卵母细胞可排卵,排卵期仅为400~500。青春期后,原始卵泡经过连续的发育、分化和成熟,直至成熟的GRAFAI卵泡。卵泡然后破裂,释放卵子。卵泡破裂后的黄体化使黄体形成。
在大约8至10周的胎儿发育过程中,卵母细胞逐渐由前体颗粒细胞包围,然后由基底层将其自身从底层基质分离出来。这种卵母细胞颗粒细胞复合物被称为原始卵泡。滤泡细胞呈立方形,滤泡周围的基质细胞突起。这一过程发生在胎儿卵巢中,在妊娠20至24周,导致初级卵泡。当颗粒细胞增殖时,透明的凝胶材料包围卵子,形成透明带。这个较大的单位称为次级卵泡。
在成年卵巢中,GRAFAI卵泡形成为最内层的三或四层快速增殖的颗粒细胞变得立方并粘附在卵子(卵丘)上。此外,在颗粒细胞之间形成充满液体的窦。随着继续积累,窦腔扩大,中心的初级卵母细胞偏心地移向卵泡壁。积云的颗粒细胞最内层与透明带紧密接触,伸长并形成放射冠。电晕放射在卵母细胞排卵时释放。覆盖颗粒细胞是一层薄的基底膜,结缔组织细胞在其外部组织成两层:鞘内层和外膜。
在每个周期中,招募一组卵泡用于发育。在许多发育的卵泡中,只有一个(优势卵泡)通常继续分化和成熟成排卵的卵泡。剩下的卵泡闭锁。在体外测量局部类固醇水平的基础上,生长卵泡可以分为雌激素主导型或雄激素主导型。直径大于10 mm的卵泡通常以雌激素为主,而较小的卵泡通常以雄激素为主。成熟排卵前卵泡的平均直径约为18~25μmm。此外,在雌激素优势卵泡中,窦FSH浓度继续升高,而血清FSH水平从中期卵泡期开始下降。在较小的雄激素占优势的卵泡中,窦液FSH值降低,而血清FSH水平下降;因此,滤泡内类固醇环境似乎在确定卵泡是否成熟或闭锁中起着重要的作用。额外的卵泡可以“拯救”闭锁通过给予外源性促性腺激素。
卵泡成熟依赖于FSH和LH受体的局部发育。FSH受体存在于颗粒细胞上。FSH刺激下,颗粒细胞增生,卵泡FSH受体数量增多。因此,生长的初级卵泡对FSH刺激的敏感性越来越高,因此,雌二醇水平增加。雌激素,特别是雌二醇,增强FSH受体的诱导,并与FSH协同作用以增加LH受体。
在卵泡发生的早期阶段,LH受体仅存在于卵泡膜内层。LH刺激诱导甾体发生,并增加雄激素合成的卵泡膜细胞。在非优势卵泡中,高局部雄激素水平可增强卵泡闭锁。FSH诱导卵泡形成,在颗粒细胞内形成芳香化酶及其受体。结果,在优势卵泡膜内层中产生的雄激素扩散到颗粒细胞中,并被芳香化为雌激素。FSH也增强了卵泡颗粒细胞中LH受体的诱导,这是注定排卵的。这些对于LH激增的适当反应是必不可少的,导致成熟期、排卵期和黄体期产生亲孕酮的最后阶段。因此,更多的FSH受体和颗粒细胞的存在和芳香化酶及其受体的诱导增加可以区分最初排卵的卵泡最终排卵和闭锁的卵泡。
诸如胰岛素、胰岛素样生长因子(IGF)、成纤维细胞生长因子(FGF)和表皮生长因子(EGF)等生长因子也可能在卵泡发生中发挥显著的促分裂作用,包括增强对FSH的响应性。卵巢肽激素,抑制素-A,抑制素-B,和激活素,在促性腺激素调节中起作用。两种形式的抑制素抑制FSH,而激活素刺激FSH释放并增强其在卵巢中的作用。卵巢颗粒细胞也产生Mü勒抑制激素(MIH),其水平现在提供了更准确的评估卵巢储备和潜在的女性生育能力(见第34章)。
排卵
排卵前LH刺激引发一系列结构和生化改变,最终导致排卵。 在排卵之前,发生整个毛囊壁的整体溶解,特别是卵巢表面的部分。 推测这是由于蛋白水解酶的作用而发生的。 随着表面细胞变性,形成柱头,并且滤泡基底膜最终通过柱头膨胀。 当这种破裂时,卵母细胞连同放射状的冠状细胞和一些卵丘细胞被排入腹膜腔,并发生排卵。
排卵是现在已知的超声研究是一个渐进的现象,卵泡的崩解从几分钟到长达一个小时或更长时间。卵母细胞附着在卵巢表面,允许延长输卵管的肌肉收缩,使输卵管上皮与输卵管上皮接触。可能肌肉收缩和输卵管纤毛运动有助于卵母细胞进入,并沿着输卵管运输。纤毛活动可能不是必需的,因为一些动纤毛的妇女也会怀孕。
初生卵母细胞处于第一减数分裂前期(双线期)。它们继续在这个阶段,直到下一个成熟分裂发生(几年后)与中环LH蜂拥而来相结合。排卵前几个小时,染色质被分解成不同的染色体,减数分裂发生在细胞质的不均匀分布,形成次级卵母细胞和第一极体。每个元素包含23个Ch-染色体,每个都是两个单子的形式。第二成熟纺锤体立即形成,卵母细胞留在卵巢表面。在排卵和受精发生后,没有进一步的发育。那时,在雄性和雌性原核的结合之前,另一个分裂发生将卵原核的染色体组分减少到23个单染色体(22加X或Y),每一个染色体由一个单子组成。由此形成卵子和第二极体。第一个极体也可以分裂。
黄体生成与黄体功能
排卵后,在LH的影响下,破裂卵泡的颗粒细胞经历黄体化。这些黄体化颗粒细胞,加上周围的卵泡膜细胞,毛细血管和结缔组织,形成黄体,产生大量的孕酮和一些雌二醇。黄体的正常功能寿命约为9至10天。这一次,它回归,除非怀孕发生,月经随之,黄体逐渐被称为白色念珠菌的无血管疤痕取代。在一个完整的周期中发生在卵巢中的事件如图4-6所示。
子宫内膜的组织病理学
子宫内膜对循环的孕激素、雄激素和雌激素有独特的反应。正是这种反应性导致月经,并使植入和怀孕成为可能。
在功能上,子宫内膜被分为两个区域:(1)外周或功能性,在月经周期中经历形态和功能的周期性变化,在男性收缩时脱落;(2)维持不变的内部或基底。每个月经周期和月经后,提供干细胞更新功能。基底动脉是基底动脉中发现的规则血管,而螺旋动脉是功能性血管中见到的特别卷曲的血管。
子宫内膜组织的周期性变化可分为月经期、增殖期或雌激素期、分泌期或孕期三个阶段。

图4-6完整卵泡周期期间卵巢发生的事件序列的示意图。 (Adapted from Yen SC, Jaffe R, editors: Reproductive endocrinology, Philadelphia, 1978, Saunders.)
月经期
因为它是外周唯一可见的一部分,所以月经的第一天是月经周期的第1天。月经的前4到5天定义为月经期。在这一阶段,子宫内膜腺和基质、白细胞浸润和红细胞外渗有破裂和崩解。除了这种功能性的剥离,由于地下物质的损失,基底的压缩。尽管有这些退行性改变,但在子宫内膜的基底层通常出现早期组织再生的早期证据。
增生期
增生期的特征是子宫内膜增殖或继发于雌激素刺激的生长。 由于子宫内膜腺的基部位于基底深处,这些上皮细胞在月经期间不被破坏。
在该周期的这个阶段,雌激素分泌的大量增加引起上皮内层里,子宫内膜腺体和基质结缔组织的显著细胞增殖(图4-7)。 在这些组织中存在许多有丝分裂,并且螺旋动脉的长度增加,其几乎遍布子宫内膜的整个厚度。 到增殖期结束时,细胞增殖和子宫内膜生长达到最大值,螺旋动脉伸长和卷曲,子宫内膜腺体是直的,狭窄的管腔含有一些糖原。
扩大腺体

图4-7早期增生期子宫内膜。 请注意由假复层柱状细胞排列的规则管状腺体。
分部阶段
排卵后,黄体分泌黄体酮刺激腺细胞分泌糖原,粘液和其他物质。 腺体变得弯曲,管腔扩张并充满了这些物质。 基质变得水肿。 有丝分裂是罕见的。 螺旋动脉继续延伸到内中膜的浅层并变得复杂(图4-8)。
腺体扩大

图4-8晚期分泌期子宫内膜。 请注意流明中分泌物的子宫内膜腺的扭曲,锯齿状外观。 该阶段的基质水肿和坏死,导致月经时子宫内膜脱落。
子宫内膜组织学在分泌期发生的显著变化允许分泌性子宫内膜的相对精确的定时(测年)。
如果在第23天没有发生妊娠,黄体开始消退,黄体酮和雌二醇分泌减少,子宫内膜退化。 在男子结构发病前约1天,发生螺旋小动脉的明显收缩,引起子宫内膜局部缺血,随后发生白细胞浸润和红细胞外渗。 据认为,这些事件发生在子宫内膜前列腺素产生的第二次。 由此产生的坏死导致月经或子宫内膜脱落。 具有讽刺意味的是,月经周期在临床上标志着月经周期的开始,实际上是生理过程的终极事件,使得子宫可以被预先准备接受另一个概念。 表4-1总结了“综合”女性生殖周期的四个组成部分。
精子发生,精子获能和受精
受精或受孕是男性和女性原核元素的结合。 受孕通常发生在输卵管中,之后受精卵继续进入子宫,发生着床并继续发育概念。
精子发生需要大约74天。 与运输一起,精子射精前总共约3个月。 精子在通过附睾期间获得动力,但精子获能使其能够在体内受精,直到它们在射精后从精浆中除去才会发生。 有趣的是,从附睾和睾丸抽出的精子可以用于体外受精,采用细胞质内注射技术直接进入卵质。
在排卵时雌激素水平高,导致数量增加,粘度降低和宫颈粘液的有利电解质含量。 这些是精子穿透的理想特征。 平均精液含有2至5毫升的精液; 40至3亿精子可能沉积在阴道内,其中50-90%是形态正常的。 不到200个精子接近卵子。 只有一个精子会在排卵时释放一个卵子。
精子的主要损失发生在阴道随后的性交过程中,从精液中排出精液起着重要作用。 另外,通过阴道酶消化精子,通过阴道酸度破坏,沿着生殖道吞噬精子,以及进一步通过输卵管进入腹膜腔的损失都会减少能够达到受精的精子数量。
那些从精子的碱性环境迁移到从宫颈口渗出的宫颈粘液的碱性环境的精子沿着粘稠度较低的粘液通道进入宫颈隐窝,在那里它们被储存以备后来上升。 可能会发生两次通向管道的波浪。
子宫收缩可能通过精浆中的前列腺素促进,在5分钟内推动精子进入管道。 一些证据表明,这些精子可能不如那些晚些时候主要依靠自己的力量到达的精子。 精子可能长期存在于腹膜腔内,但不知道它们是否具有受精能力。 Ova通常在排卵后12小时内受精。
获能是精子在受精前必须在女性生殖道中经历的生理变化。 人类精子也可以在确定的培养基中短暂孵化后获得能力,而不会留在女性生殖道中,这可以进行体外受精(见第34章)。
顶体反应是获能的主要组成部分之一。顶体,一种修饰的溶酶体,位于精子头部,作为一种“化学钻头”,旨在使精子能够进入卵母细胞(图4-9)。上覆的质膜变得不稳定并最终分解,释放透明质酸酶,神经氨酸酶和电晕分散酶。束缚于剩余的顶体顶膜的顶体蛋白可能在透明带的最终穿透中起作用。后者含有质膜特异性的质膜受体。穿过透明带后,精子头部的后额叶皮肤区域与卵母细胞膜融合,精子核结合到卵质内。该过程触发释放位于卵母细胞周围的皮质颗粒的内容物。这种皮质反应导致卵母细胞膜和透明带的变化,阻止进一步的精子进入卵母细胞。

图4-9精子头部。
获能过程可能受到精液因素的抑制,从而保持酶的最大释放,以使卵母细胞周围的电晕和透明带有效渗透。
卵母细胞的细胞进入可进一步激活精子,从而促进卵母细胞膜的渗透。对于正常受精来说,电晕不是必需的,因为它的去除对体外受精的速率或质量没有影响。这些周围颗粒细胞及其细胞间基质的主要功能可能是黏性物质,导致粘附于卵巢表面和输卵管上皮的粘膜。
在卵母细胞穿透后,精子核解聚形成雄性原核,在雌雄原核中接近并最终融合,形成合子。受精恢复染色体的二倍体数目并决定受精卵的性别。在严重精子异常引起的不孕不育夫妇中,受精卵和随后的妊娠可以在注射有或不带尾部的单个精子进入卵母细胞的细胞质后成功实现(见第34章)。
卵裂,胚,囊胚
受精后,卵裂发生。这是由一系列快速的有丝分裂分裂产生桑椹状肿块,称为桑椹胚。流体是由桑椹胚的外细胞分泌的,一个单一的充满液体的腔发育,称为囊胚腔。可以定义内部细胞团,偏心地附着于扁平细胞的外层,后者成为滋养层细胞。发育阶段的胚胎被称为胚泡,此时透明带消失。胚芽囊肿细胞可以去除和测试遗传印迹,而不损害胎体的进一步发育。
植入
受精卵在排卵后约3天到达子宫内膜腔。
激素影响卵子的运输。雌激素会导致输卵管内的“锁定”,而黄体酮会逆转这种作用。前列腺素具有不同的作用。前列腺素E舒张输卵管峡部,而前列腺素F刺激输卵管运动。卵子转运异常在不孕不育中的作用尚不清楚,但在动物实验中,卵子运输的加速导致植入失败。输卵管上皮和胚胎可释放更多的细胞因子,以促进胚胎的运输和发育,并提示即将植入子宫内膜。
最初的胚胎发育主要发生在输卵管壶腹部分,随后通过峡部快速转运。这个过程大约需要3天。在到达宫腔时,胚胎在植入前进行2至3天的进一步发育。透明带脱落,囊胚粘附在子宫内膜上,这一过程可能取决于胚胎表面特性的变化,如电荷和糖蛋白含量。各种蛋白水解酶可能在分离子宫内膜细胞和消化细胞间基质中发挥作用。
最初,胚泡面对子宫腔的壁由单层扁平细胞组成。相对厚的壁有两个区域:滋养层细胞和内细胞团(胚胎盘)。后者在7.5天内分化为厚板的原始“背”外胚层和下层的“腹”内胚层。在胚胎盘和滋养层之间出现一组小细胞。在它们体内形成一个空间,变成羊膜腔。
孕酮作用下,妊娠子宫内膜发生蜕膜改变。子宫内膜间质细胞扩增并形成多边形或圆形蜕膜细胞。细胞核变成圆形和囊泡,细胞质变得透明,稍嗜碱性,并被半透明膜包围。妊娠期蜕膜增厚5~10 mm。基底蜕膜是植入部位下方的蜕膜层。整合素是一类参与细胞间粘附的蛋白质,在植入时在子宫内膜内呈高峰,并可能起重要作用。附加的生长因子以协同的方式作用以增强植入过程。荚膜蜕膜是覆盖卵子并从子宫腔的其余部分分离出来的一层。子宫蜕膜(Pelieta)是子宫腔的剩余内层(图4-10)。囊膜蜕膜与蜕膜之间的间隔被包膜和维拉融合第四个月。
基底蜕膜进入胎盘基底板的形成。基底蜕膜海绵区主要由动脉和扩张静脉组成。基底膜蜕膜广泛被滋养细胞巨细胞侵袭,其最早出现于植入时。此时母体血清中出现HCG的微小水平。Nita -buch层是一个纤维样变性的区域,滋养层与蜕膜相遇。当蜕膜有缺陷时,如在胎盘植入物中,不存在胚层。

图4-10植入早期。
当游离囊胚在4至6天内接触子宫内膜时,合体合体滋养细胞是细胞的合体细胞,与细胞滋养层分化。在大约9天,腔隙,不规则的流体填充空间,出现在增厚滋养细胞同步囊内。紧随其后的是母体血液在腔隙内的出现,因为母体组织被破坏,母亲毛细血管的壁被侵蚀。
胎盘
由于胚泡更深地进入内胚层,滋养层束分支形成坚实的原始绒毛穿过腔隙。绒毛在受精后的第十二天首先被区分出来,是决定性胎盘的基本结构。最初位于卵的整个表面上,绒毛后来消失,除了最深的植入部分,未来的胎盘部位。
胚胎间充质首先出现在囊胚腔中的孤立细胞。当腔完全内层中胚层时,它被称为胚外细胞。它的膜,绒毛膜,由滋养层和间充质组成。当滋养层细胞被间充质核侵入时,可能来源于滋养层细胞,形成次级绒毛。
受精后约15天,母体静脉窦被抽吸。 到第17天,胎儿和母体血管都有功能,并建立胎盘循环。 当胚胎的血管与由细胞滋养层形成的绒毛膜血管相连时,胎儿循环完成。 细胞滋养母细胞在绒毛顶端的增殖产生细胞滋养层细胞,这些细胞逐渐通过外周合胞体延伸。 来自相邻绒毛柱的细胞滋养层延伸结合在一起形成细胞滋养层壳,其将绒毛附着于蜕膜。 到发育的第19天,细胞质 - 成脂细胞壳厚。 绒毛包含绒毛膜中胚层的中央核心,血管正在发育,以及合体滋养层或合胞体的外部覆盖物。
三周时,绒毛膜与蜕膜的关系明显。绒毛的大部分被绒毛剥落,被称为绒毛膜绒毛(平滑绒毛膜)。直到近第三个月的结尾,绒毛膜绒毛仍然由羊膜外腔与羊膜分离。在那里,羊膜和绒毛膜紧密接触。与基底蜕膜相邻的绒毛扩大和分支(绒毛膜绒毛)并逐渐呈现完全发育的人类胎盘的形态(图4-11)。16~20周,绒毛膜与子宫蜕膜接触并融合,从而使大部分子宫腔消失。

图4-11绒毛膜与胎盘的关系。
羊水
在正常妊娠期间,羊水腔允许胎儿室生长、运动和发育。如果没有羊水,子宫会收缩并压迫胎儿。在早孕早期羊水渗漏的情况下,胎儿可能会出现结构异常,包括面部变形、肢体缩小和继发于子宫压迫的腹壁缺损。
对于妊娠中期(20周),羊水对于胎儿肺发育变得越来越重要。后者需要充液的呼吸道和胎儿在子宫内“呼吸”的能力,使羊水流入和流出肺。妊娠中期缺乏足够的羊水与出生时肺发育不良有关,这往往与生活不相容。
羊水对胎儿也有保护作用。它含有抗菌活性,并能抑制潜在致病细菌的生长。
在分娩期间,羊水继续作为胎儿的保护介质,帮助子宫颈扩张。早产儿,其脆弱的头部,可能受益于大部分羊膜完整的分娩(en caul)。此外,羊水可以作为胎儿的沟通手段。胎儿成熟度和分娩准备就绪可通过胎儿尿液分泌到羊水中向母体子宫发出信号。
参考:hacker & moore’s essentials of obstetrics & gynecology---6e |

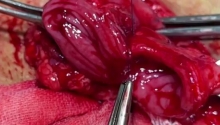
 因缺血而行小肠分段切除及一期吻合术
因缺血而行小肠分段切除及一期吻合术
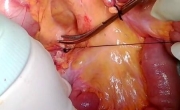
 肌肉活检 _ 如何进行肌肉活检
肌肉活检 _ 如何进行肌肉活检
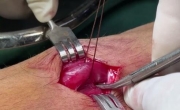
 尿道下裂修复后包皮瘘和龟头隐裂
尿道下裂修复后包皮瘘和龟头隐裂
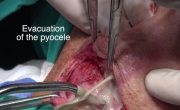 通过外部方法对喉囊肿进行有袋化或去顶
通过外部方法对喉囊肿进行有袋化或去顶

 发表于 2018-6-18 12:47:12
发表于 2018-6-18 12:47:12
 提升卡
提升卡 置顶卡
置顶卡 沉默卡
沉默卡 喧嚣卡
喧嚣卡 变色卡
变色卡 千斤顶
千斤顶 显身卡
显身卡